 Noticias de Antropologia y Arqueología
Noticias de Antropologia y Arqueología
Colonización Humana del NOA y Variación en el Consumo de los Recursos:La Ecología de los Cazadores Recolectores de la Puna Durante la Transición Pleistoceno- Holoceno. (1ra parte) |
Hernán J. Muscio
CONICET. Instituto de Ciencias Antropológicas (UBA) Sección Prehistoria. 25 de Mayo 217 4º Cap. Fed. Argentina. hmuscio@iname.com
En la Transición Pleistoceno/Holoceno la evidencia de la colonización humana del NOA, presenta en los aleros y cuevas de la Puna, una diversidad de especies potenciales como alimentos relativamente alta. En una escala regional vegetales y animales aparecen incluidos con distintas frecuencias. Los recursos de mayor rendimiento -artiodáctilos- se presentan siempre en los conjuntos, mientras que los de bajo rendimiento -roedores, aves pequeñas y vegetales- varían espacial y temporalmente. Tales variaciones aparecen en asociación con diferencias ecológicas del hábitat. A partir del análisis de una serie de modelos de densidad local de los recursos, se propone que estas variaciones arqueológicas pueden ser la señal de un juego de comportamientos de subsistencia, no siempre en fase con una "respuesta óptima" de Alto Beneficio en el largo plazo, pudiendo ser la evidencia de situaciones de abandono y retracción del rango colonizado, o de adaptaciones fallidas. Esta interpretación, privilegia las diferencias en la adaptividad potencial de la economía del nicho de caza y recolección, que habría presentado variantes No Adaptativas por situaciones de estres en la subsistencia. Estas habrían sido consecuencias de las alteraciones ecológicas del hábitat ocurridas durante la Transición. La importancia de estos registros es que pueden aportar la evidencia no esqueletal (bioarqueológica) de situaciones de maladaptación y pueden explicar el ritmo lento de la colonización humana de este bioma.
CONTENIDO
- Poblamiento Inicial del NOA y Consumo de los Recursos
- Colonización Temprana
- Escalas Espaciales y Restricciones Ambientales En la Puna
- Registro Arqueológico
- La Explicación de las Variaciones Locales del Registro Arqueológico
- Algunas Reflexiones Finales
- Comentarios de - Luis Alberto Borrero - José Luis Lanata
- Anexo
- Bibliografía Citada
Poblamiento Inicial del NOA y Consumo de los Recursos
En el ámbito de los Andes Meridionales la colonización de los diferentes ambientes del Noroeste Argentino por los humanos modernos, parece ser un proceso que comienza hacia los 11Ka BP. La evidencia arqueológica principal de este proceso proviene de las cuevas y aleros en las quebradas protegidas de la Puna, indicando una escala espacial muy amplia para la movilidad y la interacción social de las poblaciones colonizadoras, donde estarían implicados diversos ambientes tanto de las Tierras Altas como de las Tierras Bajas Subandinas. Los sitios mejor conocidos -en temporalidad radiocarbónica y en composición arqueofaunística son los de las quebradas de Inca Cueva, Huachichocana y Alero de las Circunferencias de la Provincia de Jujuy; y los de Quebrada Seca en Catamarca.
A partir de las propiedades morfológicas de los artefactos líticos, especialmente puntas y bifaces de contextos de superficie no fechados, y de la similitud con otros registros andinos, la sistemática arqueológica clásica (Cigliano 1965, 1966, Schobinger 1988) construyó un modelo para el poblamiento humano del NOA en la Transición Pleistoceno / Holoceno / Holoceno Temprano. Sin ningún soporte empírico concreto (por ej.: arqueofaunas bien fechadas), se propuso una fase inicial de poblamiento caracterizada por una tecnología de bifaces y por una subsistencia basada en la caza de mamíferos mediano/grandes y en el consumo de vegetales. Posteriormente, hacia el Holoceno Temprano se propuso una fase de cazadores especializados, con una tecnología de puntas lanceoladas. También se sugirió una movilidad moderada, siguiendo circuitos estacionales locales (fase Tuina del modelo de Nuñez y Santoro 1988). Todo esto es importante, porque en el proceso pancontinental de colonización de nuevos hábitats la tecnología, las estrategias de consumo de recursos y la movilidad fueron los aspectos claves. En este contexto, una alta movilidad y un uso oportunista de los recursos de caza, no especializado es lo defendible (ver Borrero y Franco 1997; Kelly y Todd 1988).
Más recientemente, los estudios de diversidad taxonómica de contextos fechados, realizados desde enfoques ecológicos y sistémicos, asignan una vía trófica básicamente secundaria a los cazadores recolectores tempranos de la Puna, con énfasis en la caza de camélidos (Aschero 1994), y con una estrategia de consumo oportunista de la fauna. El conjunto de la evidencia regional sustenta una hipótesis de estrategia de subsistencia con un diseño de alto beneficio -que consume principalmente biomasa animal- y manejo de la varianza -que diversifica la base de nutrientes y amortigua la disminución no previsible de los recursos (Muscio 1996, 1998). Este diseño habría resultado potencialmente adaptativo en ambientes inestables y de baja Productividad Primaria (PP). A una conclusión similar llega Yacobaccio (1994).
En general, todos los sitios de la transición Pleistoceno / Holoceno muestran que los recursos de alto rendimiento: Artiodáctilos están siempre presentes. Sin embargo, este patrón regional registra variación espacial y temporal en la inclusión de recursos de bajo rendimiento (roedores, aves pequeñas, vegetales -especialmente semillas). Notablemente, mientras Camelidae y Chinchillidae son siempre los taxa más representados en la dieta, la abundancia de cada uno varía mucho en cada depósito. Este es el caso de Inca Cueva 4 y Huachichocana E3. En el primero habría predominado el consumo de los roedores - presas de bajo rendimiento- por sobre el de los camélidos. En Huachichocana la situación es la inversa.
Desde una perspectiva evolutiva es importante no asumir a estos registros como respuestas adaptativas de las poblaciones humanas al ambiente, sin considerar antes hipótesis alternativas (O’Brien y Holland 1990,1992). Más aún si se tiene en cuenta que la hipótesis de adaptación presupone una estabilidad ambiental en escala temporal amplia, equivalente al tiempo de formación del registro arqueológico en cuestión. Para los casos que nos ocupan esta no es inferior a los 1.000 años 14C (1 Ka). La estabilidad ecológica no es la señal paleoclimática para Sudamérica en la transición Pleistoceno-Holoceno (ver Adams y Faure 1997, Clapperton 1993). En el subcontinente la transición, y hasta el Holoceno Medio, está asociada con cambios rápidos en la biota, con reorganizaciones biogeográficas y extinciones a distintos ritmos, tanto en el norte sudamericano (ver Gnecco 1995) como en Fuego Patagonia (ver Borrero 1997). Esto habría implicado cambios en la distribución y la densidad de los recursos para los humanos, y situaciones ecológicas sin correlatos actuales (Huggett 1991). La variación ecológica habría respondido a los cambios en las precipitaciones y temperaturas medias a lo largo del tiempo (Nuñez et al 1996), y probablemente en la norma de estacionalidad. Esto también podría asociarse con cambios en el patrón de circulación atmosférica (Graf 1992, McGlone et al 1993; Markgraf 1980, 1985, 1987, 1989; Markgraf y Bradbury 1982). Localmente la variación en el balance hídrico habría afectado a la productividad del ambiente y modificado el paisaje fitogeográfico. Esto último está calibrado en relación a la variación en la distribución espacial de las comunidades vegetales de la puna en las secuencias de El Aguilar y Cerro Negro, para distintos momentos del Holoceno (Markgraf 1985, Fernández et al 1991). Todas estas alteraciones habrían implicado un escenario ecológico muy dinámico con respecto a la oferta de biomasa para el consumo humano, con situaciones de estres recurrentes y de largo plazo. Este cuadro de inestabilidad ambiental para el N.O.A, es coherente con el patrón de fluctuaciones de escala global para la Transición (Sherratt 1997).
Colonización Temprana
La colonización de los distintos biomas del planeta por los humanos modernos fue un proceso relativamente rápido en la historia evolutiva de la especie. (Cavalli- Sforza 1997, Gamble 1993, 1994), y es un producto de la dispersión global de Homo sp. probablemente posterior al reemplazo de Neanderthal (ver Tattersall 1997). En este esquema los ambientes marginales habrían sido los últimos en colonizar. Principalmente en razón de la inestabilidad ambiental y de la baja productividad ecológica (Mandryk 1993).
Para el Noroeste Argentino el registro arqueológico indica la entrada de la especie en torno de los 11 Ka BP, marcando el inicio de la colonización humana de este ambiente con la experimentación de un nicho cazador-recolector. Esta evidencia corresponde a las cuevas y aleros de la Puna. En una escala hemicontinental el proceso de expansión del rango de Homo sp, en el largo plazo, habría estado controlado por la saturación lenta y gradual de los ambientes locales de mayor PP, sin implicar procesos dependientes de la densidad de escala regional. El ritmo de este proceso habría dependido de la diversidad ecológica de las américas (Steele et al 1998).
Para el N.O A, el área ubicada por debajo del Altiplano de los Andes Meridionales, la hipótesis de traza es: Norte ® Sur / Este ® Oeste para el límite Pleistoceno-Holoceno. Esta hipótesis implica una barrera biogeográfica en los Andes y poblaciones humanas alopátricas a uno y otro lado de la cordillera. Por lo menos durante los últimos milenios del Tardiglacial, cuando la productividad de la Puna Alta era bajísima como para sostener una red reproductiva de escala regional. Esto permite la ocurrencia de procesos de divergencia evolutiva. Luego de establecidas las condiciones holocénicas el flujo habría sido multidireccional.
El ritmo lento de la colonización habría estado regulado por una estasis demográfica temprana, en una escala espacio-temporal amplia (1) durante el final del Pleistoceno y el Holoceno Temprano, y discontinuidades poblacionales o adaptaciones fallidas, de las cuales parece haber evidencia arqueológica (Muscio 1998a).
A partir de la heterogeneidad ecológica del NOA, que ordena distintos biomas (2) y ecotonos (3) en un gradiente altitudinal, se propone un modelo de dispersión de poblaciones locales, en torno a las concentraciones de recursos dentro de biomas diferenciados, y a lo largo de ecotonos (ver Butzer 1985 pp 215). Con respecto a las Tierras Altas del NOA, de este modelo se infiere que las poblaciones colonizadoras deben haber sido fracciones de una red reproductiva y de una población biológica espacialmente definida en una escala mucho más grande. La misma puede ser mínimamente restringida al Noroeste Argentino, en el ámbito de los Andes Meridionales. En este planteo el Borde de Oriental habría constituido una franja longitudinal de ocupación, con ambientes de ecotono entre los ecosistemas de desierto de altura y aquellos de mayor Productividad Primaria de los Valles Mesotermales. En este paisaje, las quebradas transversales constituyen los elementos geomorfológicos más importantes como vías de acceso a la Puna. En consecuencia una implicancia importante de este modelo, es la predicción de una interacción social de los cazadores recolectores en un espacio social de escala muy amplia.
En el proceso de colonización, un modelo de comportamiento social flexible con un acceso recíproco a los ambientes locales, y la circulación sin restricciones de recursos, información y genes es lo esperable. Esto explicaría el llenado del espacio mediante la segregación de fracciones de población (ver Hammel y Howel 1987), por núcleos de población de mayor escala en los valles de menor altitud. Esto podría asociarse tanto con las escalas de las redes de intercambio de recursos (por ejemplo vegetales comestibles o materias primas líticas), como con los patrones de distribución de los diseños de arte rupestre, en un espacio social y biológico de escala muy amplia, inclusiva de la Puna.
Con fundamento en la teoría ecológica, puede sostenerse un patrón de colonización temprano que sigue un modelo de distribución heterogénea (Margalef 1980). El modelo Gruyere (ver Rapoport y Drausal 1975) parece ser el más útil -Ilustración 1-. El mismo establece: A) Un núcleo de población compacto y más denso en el exterior del área en colonización, con una mayor intensidad de mecanismos denso- dependientes (4) y estrategas + k. Este núcleo habría sido parte de una población (metapoblación) de una escala espacial abarcativa de los Andes Meridionales. En el NOA las poblaciones más densas ocurrirían en los ambientes valliserranos de mayor PP . B) Una segregación de demes (5) colonizadores desde los núcleos más densos hacia nuevas áreas marginales C) Un frente de avance con una menor intensidad de mecanismos denso-dependientes, con predominio en los bordes de estrategas + r.
El modelo Gruyere aplicado a la biogeografía humana del N.O.A, durante la Transición, implica que hacia los bordes de las áreas más marginales de la distribución de poblaciones, en las Tierras Altas del NOA, son frecuentes las retracciones por el abandono del hábitat y las colonizaciones fallidas (procesos de sístole y diástole de la expansión).
El trabajo que sigue se orienta a determinar las señales arqueológicas de estos procesos de retracción del rango colonizado, en la conducta de subsistencia. En una escala pancontinental puede defenderse un modelo de subsistencia y dispersión, basado en la caza oportunista de mamíferos y en la expansión de los rangos de caza (Borrero 1988, 1990 Kelly y Tood 1988). Esta conducta tendría su contraparte en el comportamiento tecnológico (ver Borrero y Franco 1997, Tankersley 1998), con probables variaciones clinales en los artefactos (Tankersley 1994).
El modelo que aquí se sostiene también propone un uso extenso de la biomasa animal en la dieta, controlado por la disponibilidad y la diversidad local. Las variaciones locales en el consumo de los recursos, presentes en la evidencia arqueológica de las cuevas y los aleros de la Puna, apoyan esta proposición.
En el Alero de las Circunferencias el patrón bimodal de variación en la frecuencia de arqueofaunas ocurre temporalmente: con picos en las frecuencias de camélidos y chinchíllidos en bloques radiocarbónicos discretos. Este sitio, localizado en una cota de 3600 m/nm presenta en la Capa 6 extracción 2 un dato radiocarbónico de 9.180 ± 230 AP.
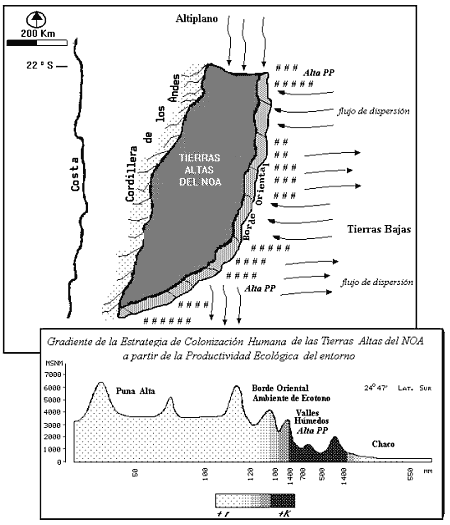
Ilustración 1: Dispersión de las poblaciones de cazadores recolectores en el NOA siguiendo la distribución de la Productividad Primaria y a partir de un modelo Gruyere. Durante el Tardiglacial la Cordillera de los Andes habría funcionado como barrera biogeográfica para el flujo humano. Los ecotonos habrían sido los ambientes preferenciales de ocupación, la Puna Alta habría sido el área de menor densidad demográfica y mayor discontinuidad poblacional.
En la secuencia de ocupación del Alero de las Circunferencias , Elkin (1994) encuentra un alto predominio de roedores por sobre artiodáctilos en las ocupaciones tempranas del alero. En el sitio los especímenes de camélidos habrían correspondido a Lama guanicoe, y en la extracción 1 de la misma capa se identificaron especímenes de taruca (Hippocamelus antisensis). La distribución estratigráfica de las arqueofaunas es de la siguiente manera:
« Para las ocupaciones arqueológicas anteriores a ca de 9.200 AP los roedores parecen haber tenido una importancia económica mayor a la de artiodáctilos tales como camélidos, aún teniendo en cuenta las diferencias existentes en el rendimiento económico respectivo. Hacia el 9200 AP la importancia económica relativa de los roedores decrece y es reemplazada por un consumo más intensivo de camélidos, para todas las ocupaciones consideradas es posible que ciertas especies de aves hayan constituido un recurso económico complementario » (Elkin 1994:148-149).
En otro trabajo se sugirió que estas variaciones en el consumo de los recursos pueden ser interpretadas como diferencias en la adaptividad potencial (6) de los comportamientos de subsistencia, y como probable señal arqueológica de rasgos no adaptativos de la economía del nicho cazador recolector durante la colonización humana de la puna (Muscio 1998). En consecuencia serían la evidencia de los procesos de retracción del rango en colonización que postula el modelo de dispersión para estos ambientes del NOA. La Ilustración 2 modela esta interpretación a partir de las arqueofaunas del Alero de las Circunferencias. La hipótesis, siguiendo el criterio de optimización, implica que la adaptividad potencial en el nivel del individuo es una función del rendimiento promedio de la dieta.
Los roedores, que aparecen dominando la composición de la dieta son recursos de muy bajo rendimiento económico en relación a los artiodáctilos. En el gráfico esto está expresado como un cambio en la adaptividad del componente de subsistencia del nicho, que se eleva hacia un pico explicado por el beneficio ganado como consecuencia de un alto consumo de los artiodáctilos. En el caso de Inca Cueva y Huachichocana esta variación ocurre espacialmente, como variantes locales dentro de un lapso radiocarbónico comparable. En cambio en Quebrada Seca 3, el consumo de carne de camélidos se mantuvo dominando la dieta, y de una manera más o menos homogénea, durante todo el Holoceno Temprano.
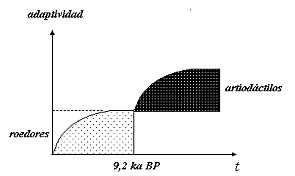
Ilustración 2: Modelo de la variación en la adaptividad potencial del consumo humano de recursos, durante el Holoceno Temprano en el NOA, a partir del registro del Alero de las Circunferencias.
Este artículo, en el marco de la Arqueología Evolutiva (Lanata 1999), analiza estas variantes en el consumo de los recursos en términos ecológico-evolutivos, esto es: considerando que las poblaciones humanas forman parte de ecosistemas dinámicos. El objetivo básico es la búsqueda de hipótesis alternativas a la de adaptación, para comenzar con el desarrollo de un escenario biogeográfico y evolutivo para los cazadores recolectores del NOA (ver Lanata y Neff 1998 MS), que incluye probables situaciones de adaptación fallida o retracción. La metodología prioriza el estudio de la variabilidad del registro de la Puna, observando la manera en que se aparta de una respuesta óptima. Siguiendo la línea de trabajos anteriores (Muscio 1996, 1998), interesa construir herramientas teóricas para discriminar de la flexibilidad del comportamiento de subsistencia -propia de la especie- probables señales arqueológicas tanto de nichos más o menos exitosos, como de maladaptación (ver Borrero 1993).
Escalas Espaciales y Restricciones Ambientales en la Puna
Partiendo de la estructura ecológica de la Puna y considerando las propiedades de los recursos, es posible derivar hipótesis acerca de la adaptividad potencial de los comportamientos de subsistencia puestos en práctica durante la colonización humana en el Tardiglacial. Para esto se utilizan criterios de optimización, a partir de restricciones, contextos y objetivos de optimización. (Foley 1985, 1989,1996 Stephen y Krebs 1986).
La Puna argentina es un bioma de desierto de tierras altas (por encima de los 3.000 m/nm) de bajísima productividad primaria, con un promedio teórico de ± 53 g/m2/yr (Muscio 1996, 1998b); una distribución espacial heterogénea de los recursos (en riqueza y densidad) que varía altitudinalmente, una alta estacionalidad; y una muy alta variabilidad de los elementos del clima (Muscio 1998b). En el ámbito del Noroeste argentino, hoy mismo es una región no óptima, o marginal. Esto es importante para pensar los procesos de ocupación abandono y reocupación de hábitats controlados por las variaciones físicas del paisaje, especialmente por episodios de sequía severa (ver Avery 1995). Se puede plantear que en este ambiente la escasez y la irregularidad de la oferta de nutrientes controlan los principales estresores no dependientes de la densidad de la población, para un nicho cazador-recolector.
Es importante señalar las escalas espaciales en las que se evalúan las conductas de subsistencia a partir de un perfil óptimo. Teniendo en cuenta una población humana espacialmente continua (Wobst 1993) y desde una perspectiva ecológica, se puede postular que existirán variaciones geográficas correlacionadas con las propiedades de los ambientes locales. En este trabajo se considera al bioma como el componente espacial de escala macroregional - que en este caso comprende al desierto de altura de la Puna. Dentro del ambiente regional amplio, se trabaja con ambientes locales resueltos entre los 10 Km2 y los 300 Km2 (Muscio 1998b). La escala local capta ambientes que pueden ir desde un sistema de "vega" o una quebrada, hasta un valle con distintos mosaicos de recursos para el hábitat de poblaciones humanas locales. Se deduce que una conducta de alto rendimiento en su entorno local -localmente óptima - puede ser de beneficio parcial -subóptima- o incluso no adaptativa considerando las propiedades del ambiente y la población de escala regional. Esto también es importante para distinguir a los procesos de saturación local del ambiente, de los regionales. Siendo ambos procesos denso-dependientes, los primeros resultan del impacto de la predación humana en la base local de recursos y gatillan procesos de abandono del hábitat sin requerir una alta densidad de población regional ni competencia; mientras que los segundos sí están asociados a la densidad de humanos de gran escala espacial, y controlan la superposición y la competencia.
A partir de estas consideraciones, en el ANEXO, este trabajo fundamenta un conjunto de hipótesis de optimización, considerando diversos contextos locales como hábitats para una ocupación humana regional hipotética. A tal fin se manejan cuantitativamente el rendimiento y la densidad de los recursos. Mediante estimaciones de biomasa se construyen series locales de densidad de los recursos, para cotejarlas con su rendimiento económico potencial.
A continuación se pasa a discutir algunas propiedades del registro arqueológico de los cazadores recolectores de la Puna, y se remite al ANEXO para examinar del desarrollo de las hipótesis de optimización.
Registro Arqueológico
Esta sección analiza la evidencia arqueológica de la Puna a la luz del modelo de estrategia general de dispersión y colonización del N.O.A. Principalmente se focaliza en dos aspectos del registro: diversidad arqueofaunística y localización espacial del hábitat. Los datos manejados son principalmente de 2 sitios: La cueva III de Huachichocana (CH III) capa E3, e Inca Cueva 4 (ICc4) capa 2. Ambos sitios se emplazan en el Borde Oriental de la Puna jujeña entre los 3.400 y 3.600 m/nm. y tienen acceso a una alta diversidad de ambientes localizados por encima y por debajo de esta cota. Se prioriza estos sitios dado que tienen controles tafonómicos. Especialmente en relación a la acción de carnívoros afectando la integridad y la representación de las arqueofaunas. El bloque temporal para estas ocupaciones está entre los 10,6 y los 7,4 Ka BP (Aschero 1984, Fernández Distel 1974, 1986). También se mencionan algunas propiedades del registro de Quebrada Seca (QS3) en Catamarca.
Movilidad y Escala Espacial del Nicho: Los registros de CH III e ICcE, sustentan la hipótesis de alta movilidad, indicando un rango de acción extenso en una red de interacción biológica y social de gran escala. La evidencia incluye elementos subandinos (Aschero 1975, Fernández Distel 1974, Yacobaccio 1983, 1985). Las plumas de aves tropicales Ara militaris, están envueltas por un cordel o vena muy fina (Yacobaccio 1985). También hay cuentas de columela de Strophochelius (Megalobulimulus oblongus) y Diplodom sp, que actualmente se encuentran al este de la Quebrada de Humahuaca, en ambientes como la cuenca del río San Francisco (Yacobaccio 1983). Estos elementos habrían formado parte de artefactos suntuarios.
La procedencia de los vegetales es una evidencia de la movilidad entre los valles intermedios, las Tierras Bajas y la Puna Alta, indicando el uso de distintos hábitats locales. Principalmente los algarrobos, porotos y ajíes de Huachichocana. Lo mismo se puede sostener a partir de la procedencia de algunas materias primas líticas de Inca Cueva (Ratto 1989). Todas estas evidencias favorecen el modelo de dispersión de poblaciones humanas en torno de las concentraciones de recursos y a lo largo de ecotonos. Especialmente los de la interface ecológica Prepuna/Valles Mesotermales.
El uso de la Puna parece haber sido discontinuo a lo largo del ciclo anual, lo que se deduce del perfil etario de los camélidos consumidos y de la época de floración de los vegetales. Esto favorece la hipótesis de un patrón estacional en la movilidad, funcionando para amortiguar la caída estacional de la disponibilidad de los recursos, con la ocupación del Borde de Puna y la incursión a la Puna Alta al comienzo de la temporada húmeda (ver Yacobaccio 1991). La escala del rango anual con una movilidad estacional que abandona la Puna durante el invierno para explotar los ambientes de los valles y la quebradas mesotermales depende de la geografía local.
La evidencia de ICc4 y CH III demarca un espacio de interacción social y biológica de una escala del orden de los 5.000 Km2. Las ocupaciones de Quebrada Seca podrían implicar un rango mas extenso. Debido a que desde aquí las áreas más cercanas que pueden ofrecer hábitats comparablemente menos duros durante el invierno, se localizan a una distancia mínima de 100 Km. hacia el oeste. Si tomamos en cuenta los 70 Km. como indica la procedencia de las obsidianas para las ocupaciones de cazadores recolectores (Pintar 1992) se puede estimar un rango anual ± 25.500 Km2. Ejemplos etnográficos muestran cazadores recolectores de biomas de muy baja productividad y alta estacionalidad, con rangos anuales que superan en mucho estas estimaciones (7). Toda esta evidencia apoya la hipótesis de una población colonizadora de la Puna de escala amplia, con núcleos más densos en los valles del Noroeste Argentino.
Consumo de los Recursos: Tomando los recursos animales, Huachichocana para un total NISP de 859 elementos muestra una composición donde Chinchillidae constituye el 11,42 %, Cervidae el 0,23 % y Camelidae el 87,41 %, con una notable presencia de ejemplares inmaduros (Yacobaccio 1991, 1994). Inversamente en Inca Cueva 4 los chinchíllidos, particularmente Lagidium viscacia, son los recursos que predominan en la muestra con el 70,97 % de un total de 1045 NISP; los camélidos solo están presentes con 10,24 %, en su mayoría pertenecientes a individuos adultos y los cérvidos solo con 1,43 %.
Considerando la naturaleza promediada del registro arqueológico, interesa observar como varió en intensidad la entrada de los recursos en la dieta. Existen distintos índices que describen las variaciones en la dieta de largo plazo. Principalmente aquellos que comparan artiodáctilos vs. recursos de bajo ranking (Janetsky 1997, Spielmann y Angstadt-Leto 1996). Las tasas de descarte y depositación son indicadores adecuados de las variaciones en la frecuencia promedio de los comportamientos prehistóricos (Lanata 1993). Aquí se utiliza la Tasa de Depositación (TD) aplicada a la biomasa animal, que viene dada por el cociente:
TDa= na /t1-t2
Donde: na es la cantidad total de una clase "a" de recursos
t1-t2 es el intervalo radiocarbónico entre el dato más antiguo t1, y el más reciente t2
En una importante contribución Yacobaccio (1994) transformó las frecuencias óseas de ICc4 y CH III en números de biomasa. En el cálculo que aquí se realiza de TD se utilizan estos valores de biomasa. Por lo tanto la variable medida es la tasa de depositación de la biomasa animal, o la densidad temporal de la carne consumida. El valor de la unidad para esta variable es Kg./Ka. Los extremos radiocarbónicos del bloque temporal de ocupación de CH III son de 8,67 Ka BP / 10,20 Ka BP con una extensión de 1,53 Ka. Para ICc4 C2 estos son de 9,23 Ka BP / 10,62 Ka BP, abarcando 1,39 Ka . Estos bloques radiocarbónicos son las unidades temporales de observación de variantes en la dieta. La tabla 3 muestra la TD calculada para cada uno de los depósitos.
|
Taxa |
TD Inca Cueva 4 C2 Kg./Ka |
TD Huachichocana E3 Kg/Ka |
|
Camélidos |
14,13 |
44,87 |
Tabla 3: Tasa de depositación (DT) de la biomasa animal, en Kilogramos (KG) por Kiloaños (Ka), calculadas para ICc4 y CH III E3.
Observamos en la tabla que ambos depósitos para bloques temporales de extensión comparables muestran similares cantidades de biomasa animal consumida. La poca cantidad de este valor, un promedio de 45, 85 Kg. de biomasa animal consumida cada mil años radiocarbónicos es un indicador de la baja intensidad del uso del hábitat local. La variación discutida también está presente en la diversidad de especies consumidas -tabla 4. Paralelamente a los animales, ambos conjuntos incluyen recursos vegetales –frutos , semillas y floraciones- comestibles. Sobre una riqueza total de 18 taxones Huachichocana registra 9, mientras Inca Cueva 15.
La interpretación aquí ofrecida del registro promediado es coherente con un modelo de llenado lento del espacio por discontinuidades en las ocupaciones de ambientes locales, y puede también señalar una etapa de experimentación (sensu Borrero 1995) en la colonización de la Puna.
|
TAXON Artiodactyla |
IC 4 X |
CH III X |
TAXON Dasipodidae |
IC 4 X |
CH III - |
Tabla 4: Diversidad Taxonómica de los Registros de ICc4 y CH III E3
La Explicación de las Variaciones Locales del Registro Arqueológico
Un aspecto central en los programas de investigación sistémicos o ecológico-sistémicos fue la explicación de la variabilidad arqueológica mediante modelos de funcionalidad de sitios y de patrones de asentamiento. Sin embargo la naturaleza promediada del registro arqueológico, que está en la escala temporal de su formación, y el reconocimiento de las innumerables variaciones que pueden ocurrir en la evolución del uso humano de un paisaje, obligan a abandonar estos enfoques.
Una vía satisfactoria para la explicación de la variabilidad arqueológica en los indicadores de subsistencia del registro, es considerar a la diversidad de situaciones puntuales, como variaciones locales del consumo de los recursos en la ocupación humana regional de un lapso temporal de escala amplia. El uso promediado de los sitios permite interpretar a la riqueza y a la frecuencia de las arqueofaunas, como producto de la frecuencia de entrada de los recursos en la dieta, en una amplitud máxima, para el bloque temporal de formación del depósito (Broughton y Grayson 1993). Bajo esta perspectiva las variaciones en las tasas de depositación de arqueofaunas pueden interpretarse como variaciones en las frecuencias de consumo de recursos, bajo situaciones ecológicas y demográficas particulares.
Para los casos que venimos analizando, la evidencia arqueológica indica en Inca Cueva 4 C 2 una ocupación promediada con una alto consumo de fauna de bajo rendimiento y una alta diversidad de recursos, mientras que CH III E3 habría explotado una menor diversidad de recursos y con un alto índice de consumo de fauna de alto rendimiento. Es importante señalar que esta variaciones en la selectividad de presas no reflejan un desvío subóptimo. Los chinchíllidos no son recurso subóptimos, son de bajo rendimiento.(8) Entonces se puede postular que habrían existido diferencias de adaptividad en el comportamiento de subsistencia. Considerando los modelos de las series locales de abundancia de recursos para los distintos entornos, presentados en el ANEXO, estas variantes pueden explicarse postulando variaciones en la disponibilidad local de la biomasa.
La situación de CH III E3 tiene correspondencia con la serie teórica "óptima" de Alto Beneficio de la figura 3 del ANEXO, En la estrategia de consumo de CH III el 99 % de la carne es de alto ranking, con probables inclusiones subóptimas como los frutos de Passiflora y T. pasacana. En la composición de la dieta los vegetales representan el 30 % de la riqueza de recursos, tomando a los camélidos como recurso combinado. Esta diversificación puede amortiguar la inestabilidad ambiental sin interferir con la estrategia global de alto beneficio.
En el otro extremo de la geografía regional, en la Puna Alta de Catamarca, el registro de Quebrada Seca localizado a 4050 m/nm, también muestra un consumo de recursos en fase con lo "óptimo". El registro muestra un consumo promediado con una altísima frecuencia de artiodáctilos. Este ambiente es mucho más árido que el Borde de Puna y con un régimen de menor PP (entre los 28 y los 35 g/m2/yr, ver Muscio 1996). El entorno es de una baja densidad y diversidad biológica, lo que implica un ambiente local con mayor direccionalidad para los comportamientos de alta eficiencia en la captura y manejo de los recursos. En este ambiente la vicuña y el guanaco son los únicos ungulados silvestres de alto rendimiento, dado que aquí taruca no superpone el rango con estos taxa.
El Alero 3 (QS3) capa 2b 12-25 en el intervalo radiocarbónico 9,05 / 8,33 Ka BP para un total de 817 NISP presenta un 84,08 % de camélidos y el resto recursos de bajo ranking: chinchíllidos 9,06 % Ctenomys 0,12 % Abrocoma 0,12 % , Rodentia 4,52, anidae 0,12 y Avis 1,95 (Olivera y Elkin 1994). En estas ocupaciones el uso de los vegetales está restringido al acondicionamiento de los pisos del alero y como material combustible.
Todo esto contrasta con el registro de ICc4 C2, que se corresponde con la serie teórica de perfil de Bajo Beneficio de la figura 5 del ANEXO. La amplitud de la dieta realizada, para el lapso de formación del depósito, resulta de una alta frecuencia de consumo de recursos de bajo rendimiento: el 58 % de la biomasa animal es de esta clase. Las plumas de rheidae sugieren la caza de esta presa subóptima. En términos de riqueza de recursos, la evidencia muestra una gran amplitud -15 taxones- de los cuales 4 son recursos vegetales, incluyendo semillas, frutos y probables tallos comestibles. La alta inclusión de vegetales, especialmente semillas, se infiere de la presencia de molinos planos y manos de moler, probablemente vinculados al procesamiento de estos vegetales. La evidencia indica que esta estrategia de subsistencia incluiría con mayor frecuencia los recursos de mediano y bajo rendimiento.
En este punto es importante advertir que en la oferta regional de recursos, los de menor tamaño -vizcachas, semillas, huevos principalmente- son los de mayor riqueza de nutrientes con mucha heterogeneidad en grasas, vitaminas y proteínas (Muscio 1998). La alta inclusión de estos recursos puede estar reflejando una situación de estres en la base de nutrientes. Debido a la riqueza en proteínas, una caída regional en la disponibilidad de biomasa animal puede ser promediada con una mayor inclusión de semillas y huevos en la dieta. Los chinchíllidos y los tubérculos aportarían principalmente grasas y carbohidratos en situaciones de estres. La probable inclusión en la dieta de huevos de suri, postulable a partir de la evidencia de ICc4 C2 apoya la hipótesis de una dieta diversificada en fuentes de nutrientes.
El conjunto de toda esta evidencia sugiere un comportamiento local que desde lo óptimo no está en fase con la estrategia de Alto Beneficio (ver ANEXO), pudiendo responder a una situación de estres en la subsistencia, por escasez de recursos alimenticios de alto rendimiento. Esto podría asociarse a maladaptaciones durante el intervalo 10,6 / 9,23 Ka BP del poblamiento de la quebrada de Inca Cueva, por un comportamiento de largo plazo No Adaptativo. Esta hipótesis debe ser puesta a prueba en esqueletos humanos, buscando principalmente patologías óseas por malnutrición .
Otros registros arqueofaunísticos del Borde de la Puna pueden también ser interpretados por estas hipótesis. En la Quebrada de Pintoscayoc el Alero de las Circunferencias presentaría ambas variantes en una secuencia de ocupación. Estas variaciones tienen sentido en el modelo Gruyere de dispersión de poblaciones colonizadoras, que predice el aumento en la frecuencia de extinciones y abandono de los hábitats colonizados, conforme se avanza en las áreas más marginales de la distribución. Especialmente en estos espacios de ecotono de la prepuna, donde están localizadas las ocupaciones con un consumo de recursos de bajo rendimiento económico. De tal manera, estos registros pueden ser la señal arqueológica de adaptaciones muy costosas o quizás de adaptaciones fallidas en el Borde de la Puna, interpretables como el efecto promediado de "pulsos" de ocupación de los aleros.
Situaciones similares de estres en la subsistencia, también pudieron ocurrir en los ecotonos de la vertiente occidental de la Cordillera de los Andes, durante la Transición . Este puede ser el caso de las ocupaciones de la quebrada Intermedia de San Lorenzo, en Chile. Allí los estratos IV al IX para un rango radiocarbónico entre los 10,4 y los 9,6 Ka BP (Nuñez y Santoro 1988) presentan una composición arqueofaunística donde camelidae solo representa el 7,3 % de la muestra, predominando ampliamente los recursos de menor rendimiento: chinchillidae con el 63, 2 % , rodentia 23, 5 % y avis. 5, 8 %. A esto se le suma el consumo de vegetales, que se infiere de la presencia de "palos cavadores": artefactos específicos para la explotación de tubérculos o raíces.
Esta interpretación de la diversidad de las arqueofaunas en razón de la eficiencia económica de la estrategia de subsistencia, puede ser de utilidad junto con otras líneas de evidencia, para la discusión arqueológica de procesos de experimentación en la colonización de una región nueva para la especie. Estos procesos, que siguiendo a Borrero (1995) son de difícil identificación en el registro, implican series discontinuas de ocupación, con estrategias no óptimas y con posibles extinciones locales. Para los cazadores recolectores "colonizadores" de las Tierras Altas del NOA, todo esto apoya un modelo demográfico espacialmente muy heterogéneo, de muy baja densidad de individuos en una escala regional. Durante la Transición, y el Holoceno Temprano, la dinámica demográfica de estas poblaciones debió responder a escenarios locales de escasez de largo plazo de los recursos, los que habría inducido de manera recurrente procesos de extinción local de poblaciones, o de abandono de ambientes colonizados. A continuación se argumenta esta explicación
Alteraciones Ecológicas y Estres de la Subsistencia: La predicción básica del Modelo de la Amplitud de Dieta establece que conforme disminuye la abundancia de los recursos de alto rendimiento, aumenta la frecuencia de consumo de los recursos de bajo rendimiento. En consecuencia las variantes locales en la amplitud de la dieta de los cazadores recolectores de la Transición Pleistoceno / Holoceno, pueden interpretarse en relación a las variaciones en la disponibilidad, o en el acceso a la biomasa de herbívoros de alto rendimiento, en un contexto ecológico dinámico.
La principal característica de la ecología regional del Tardiglacial y del Holoceno Temprano, por lo menos hasta los 8 Ka BP en Sudamérica, fue la gran dinámica del paisaje vegetal, y de los procesos de rápida reorganización de la biota (ver Adams y Faure 1997, Nuñez et al 1996)). Este reconocimiento de un paisaje ecológico muy dinámico y cambiante en distintos ritmos, necesariamente requiere que se abandonen los modelos de adaptación humana de largo plazo, representados por los modelos de sistemas adaptativos en equilibrio, los cuales presuponen -contra la evidencia empírica disponible- una estabilidad ecológica en la escala temporal arqueológica (que es de resolución radiocarbónica o estratigráfica). De las características de las variaciones ocurridas en el Tardiglacial y durante el Holoceno Temprano, se deduce que los cambios en el paisaje ecológico no habrían sido modificaciones estructurales, sino alteraciones en magnitud. Las propiedades del ambiente afectadas habrían respondido al modelo de variabilidad de escala media propuesto por Butzer (1985), implicando:
«Cambios fundamentales en la hidrología, la productividad y en todas las categorías de biomasa. La composición cualitativa de las comunidades bióticas perdura, pero los cambios cuantitativos afectan a las estructuras en mosaico en general, y a los ecotonos en particular (número de especies y densidades demográficas seleccionadas)... » Butzer (1985: tabla 2-4)
La variación habría implicado fundamentalmente procesos de escala ecológica. Funcionalmente se habrían alterado las relaciones herbívoro/forraje (sensu Soriano 1987). En consecuencia los cambios en la distribución y en la productividad de la Biomasa Primaria (vegetal), resultantes de las variaciones en la temperatura y humedad deben haber alterado la disponibilidad de las principales presas de caza, fundamentalmente la abundancia de los camélidos en los ambientes locales de ecotono (9). Para las poblaciones de camélidos, el principal recurso potencial para los cazadores recolectores, es probable que la tendencia demográfica durante los períodos de disminución de nutrientes haya sido la disminución en la tasa de crecimiento, por ajuste logístico. Paralelamente, la frecuencia de los movimientos migratorios a partir de la distribución heterogénea de las pasturas debió haber aumentado, llevando a una distribución más irregular y menos densa de este recurso en el ciclo anual (10) . Estas, junto a otras variaciones de carácter ecofonotípico, serían las respuestas esperables para los ungulados en un proceso de reordenamiento de los hábitats inducido por variaciones climáticas (Vrba 1996).
En el NOA, la sucesión de comunidades vegetales en el espacio está determinada altitudinalmente, siguiendo de manera continua el gradiente de humedad del patrón de precipitaciones (Bianchi y Yañes 1992). Junto con los elementos del relieve del paisaje, esto se expresa con una alta variabilidad en el hábitat potencial para los cazadores recolectores. En el continuo ecológico, el Borde Oriental de la Puna presenta amplios ambientes de ecotono en la franja transicional entre las comunidades vegetales de las Tierras Altas de la Provincia Puneña, y la vegetación de la Provincia Prepuneña, de las quebradas y los valles de cotas inferiores a los 3000 m/nm. Estos ambientes ecotonales habrían sido los más sensibles a los efectos de las reorganizaciones biómicas (Butzer 1985) producidas por las variaciones climáticas durante la Transición. En los ecotonos las alteraciones bióticas complejas y las reorganizaciones de los ecosistemas habrían inducido a cambios en el mosaico de los recursos. Para los Andes, especialmente en la abundancia de los ungulados: camélidos y cérvidos. La estrategia de subsistencia humana debió haber sido sensitiva a los cambios en el rango de esta fauna. Principalmente los patrones de movilidad, la tecnología y la inclusión de recursos en la dieta de los cazadores recolectores (ver Pintar 1992,1994).
El reemplazo de un modelo estático del paisaje ecológico por uno más realista: altamente dinámico en múltiples escalas, permite explorar hipótesis de estres de subsistencia en el poblamiento cazador recolector, tanto en la tecnología como en la dieta realizada, en escenarios de degradación del hábitat por alteración ecológica.
Sobre esta base puede postularse que mientras los registros de Quebrada Seca 3 y Huachichocana (y quizás también las ocupaciones estratigráficamente superiores del Holoceno Temprano de Alero de las Circunferencias) serían los de grupos humanos ocupando un espacio bajo condiciones de buena disponibilidad de recursos de alto beneficio, Inca Cueva 4 C2 y el Alero de las Circunferencias pre 9,2 Ka BP, serían el producto de la ocupación humana de ambientes locales de ecotono, con una baja densidad de ungulados y con una estrategia de subsistencia bajo estres.
La secuencia de ocupación del Alero de las Circunferencias puede ser el ejemplo más claro de la variación de la amplitud de la dieta por el cambio en la ecología local de los ecotonos de la prepuna. En la secuencia del sitio las ocupaciones más antiguas, estratigráficamente las más profundas, se revelan como las de menor acceso a una dieta de alto rendimiento, lo que puede asociarse a un ambiente local poco favorable para la colonización. Luego le siguen las ocupaciones con un mejor acceso a los recursos de caza de alto rendimiento, lo que indicaría condiciones ecológicas más favorables para el establecimiento de un hábitat humano local. Patrones similares de variación en la subsistencia humana, también pudieron ocurrir durante la colonización de los ecotonos de la vertiente occidental de los Andes.
Finalmente, los contextos ecológicos alterados por las variaciones climáticas podrían favorecer, sin necesidad de una demografía alta de escala regional, procesos de extirpación de fauna por sobreexplotación, especialmente en ambientes locales de baja PP y poblaciones de ungulados poco densas y muy móviles. Todos estos procesos, darían lugar a una alta frecuencia de abandono y reocupación de ambientes locales, que se expresarían como discontinuidades en las secuencias arqueológicas, y con mucha variación local en las frecuencias taxonómicas de las arqueofaunas.
Algunas Reflexiones Finales
Mi convicción sobre el poder de la selección permanece inquebrantable; pero es probable, o casi cierto, que varias de mis conclusiones serán desechadas por erróneas con el tiempo, pues esto difícilmente deja de ocurrir siempre que se trata por primera vez un asunto cualquiera.
Charles Darwin, El Origen del Hombre
Frecuentemente la mera detección arqueológica de la presencia humana en una región y un momento sin registros previos, es considerada per se como la evidencia de una adaptación lograda y de la colonización exitosa de un nuevo hábitat. Generalmente apelando a mecanismos de ajuste a las presiones ambientales. Bajo la lógica del dispersionismo difusionista (evolución local y radiación hacia el exterior) y del adaptacionismo (los sistemas una vez evolucionados permanecen adaptados) el razonamiento está justificado. Bajo la lógica evolutiva no. Desde un punto de vista biogeográfico y evolutivo los procesos de colonización se explican considerando, principalmente la acción de la selección en la población colonizadora local, además de la deriva (ver Borrero 1993, 1995). En la perspectiva ecológico-evolutiva los procesos de extinción local, maladaptación, retracción del rango colonizado y experimentación, ocupan un lugar central en la explicación arqueológica de la colonización de nuevos hábitats para la especie. Esto necesita de los estudios de las variantes en los comportamientos y los artefactos, asociadas a las variaciones en la adaptividad potencial de la población, lo que lleva a cruzar datos con la evidencia esqueletal. Especialmente en relación a la disponibilidad y al consumo de la biomasa producida por el entorno, cuando se postula que la expansión global del rango Homo sp estuvo controlada por la saturación de los ambientes locales (ver Young y Bettinger 1995, Steele et al 1998) produciendo variantes clinales (Gamble 1993).
Este trabajo presentó un modelo de colonización del NOA considerando la dispersión de las poblaciones humanas de la interface Pleistoceno-Holoceno en la escala de los Andes Meridionales. Partiendo de criterios de optimización, sondeó en las variaciones locales de los comportamientos de subsistencia de los cazadores recolectores que colonizaron las Tierras Altas del NOA. Los casos estudiados parecen indicar patrones evolutivamente significativos. La baja intensidad de las ocupaciones sugiere un uso intermitente de los aleros y las cuevas, y comportamientos muy móviles, utilizando una amplitud grande de ambientes diversos. Esto refuta los modelos de movilidad moderada en torno a oasis o ambientes preferenciales (i.e. Cigliano 1966, Schobinger 1988), también presentados como de trashumancia inicial (Nuñez y Santoro 1988).
La evidencia regional muestra un consumo oportunista de los recursos, muy sensible a las propiedades ecológicas del entorno local. De tal manera la hipótesis de caza especializada para el Holoceno Temprano no tiene sustento en el registro arqueofaunístico. Paralelamente, se argumentó que en algunos casos las variaciones locales en las arqueofaunas, responderían a poblaciones con un consumo de recursos no en fase con lo óptimo de Alto Beneficio. Todas estas evidencias dan crédito a un modelo de llenado lento del espacio en la Puna, con una trayectoria demográfica controlada por la discontinuidad poblacional y la retracción durante el finiglacial. Esto desafía la visión adaptacionista. Con base teórica en la ecología-evolutiva se puede explorar la hipótesis según la cual, las variantes No Adaptativas de la dieta pueden ser el producto de situaciones de estres en la subsistencia, y la señal no esqueletal de maladaptación.
Respecto de la escala espacial, este trabajo sugiere que no es posible interpretar adecuadamente los procesos biogeográficos y evolutivos del poblamiento de las Tierras Altas del NOA, considerando a la Puna como un área independiente. De esto deriva la necesidad de ampliar la escala espacial de los diseños de investigación de cazadores recolectores, para comenzar a buscar la evidencia arqueológica en otros ambientes del NOA. Sobre todo en la geografía valliserrana y en las sierras subandinas. Al mismo tiempo, el conjunto de las hipótesis presentadas requiere abrir el trabajo hacia nuevas líneas de investigación. Principalmente tasas de descarte de artefactos, patrones de uso del paisaje, abandono y reocupación de aleros y cuevas; y diseño - performance de artefactos. En tal marco es importante calibrar las escalas espaciales de los movimientos de materias primas, en redes de interacción social amplias (ver Lazzari 1994). Todo esto necesita que se superen los modelos de industrias locales, realizados a partir de la representación modal de las diferentes clases de artefactos en sitios "arquetípicos", y que el registro arqueológico sea trabajado a partir de sus propiedades distribucionales (sensu Dunnel y Dancey 1983).
Un componente importante de las investigaciones futuras deben ser los estudios de las variaciones bióticas de la Transición Pleistoceno/Holoceno, focalizando en las alteraciones ecológicas con fluctuación en la disponibilidad de los recursos de los ambientes locales de caza y recolección (de una escala del orden de los 1000 Km2). Especialmente considerando que el proceso de colonización del NOA habría dependido de la dispersión por la saturación local de los ambientes explotados. En este enfoque pueden plantearse dos líneas principales de investigación, no excluyentes: A) el estudio de los movimientos y los reacomodamientos del rango de los artiodáctilos como función de las variaciones ecológicas de la interface Pleistoceno/Holoceno, comenzando por las variaciones en la productividad primaria del bioma (que lleva a la construcción de columnas paleoambientales locales). B) los procesos de saturación local de ecosistemas de baja productividad, derivados de la sobreexplotación humana (que requiere estudiar localmente el umbral de saturación del hábitat y la fauna más sensitiva de la sobrecaza). Es importante destacar, como advirtieron Kelly y Todd (1988) en el estudio de la dispersión de escala continental, que la variabilidad ecológica en la Transición Pleistoceno-Holoceno debió haber impuesto a los humanos escenarios muy diferentes a los que la etnografía de los cazadores recolectores presenta.
La perspectiva evolutiva plantea interrogantes novedosos en la arqueología del N.O.A que pueden integrar, en un marco teórico que privilegia la diversidad, el conocimiento ganado para el área bajo diferentes perspectivas. En este dirección la arqueología de cazadores recolectores del N.O.A requiere que se integren distintas vías metodológicas, adecuadas a la investigación tanto de los aspectos ecológicos de las poblaciones prehistóricas, como de los aspectos sociales (sensu Minegal 1997, ver Ingold 1993). Lo mismo puede plantearse para procesos de Evolución Social (sensu Nielsen 1995). Este desafío implica comenzar a explorar con preguntas nuevas en los patrones de la variabilidad artefactual que la sistemática clásica de la arqueología del NOA dejó al descubierto, y en la variabilidad ecológica que los enfoques sistémicos procesuales demostraron. Este trabajo buscó contribuir en el desarrollo de esta tarea.
Agradecimientos: A Luis Alberto Borrero por la lectura y la corrección de una versión previa de este trabajo y por las importantes sugerencias brindadas en la discusión del mismo (las cuales validaron la cita de Darwin). A José Luis Lanata por su lectura crítica de un manuscrito y por estimularme para su publicación. A Rolando R. Lillo por su apoyo a mi trabajo y por mejorar la elegancia matemática de cada una de mis ideas. A Mónica Ferraro, María Eugenia Cantero y al personal de la Biblioteca del Museo Etnográfico J. B Ambrossetti por su ayuda continua en la búsqueda de bibliografía. Estas menciones no implican responsabilidad alguna en las expresiones aquí vertidas.
Comentarios
NOTAS
- La estasis propuesta es un fenómeno resuelto en una escala temporal del orden de los 10 Ka, representado como la parte plana de una curva de evolución demográfica que comienza a crecer exponencialmente en torno a los 5 Ka BP. Su escala espacial mínimamente abarca el NOA (Muscio 1996).
- El concepto de Bioma refiere a una comunidad de animales y plantas que ocupan un área climáticamente uniforme en una escala continental (Hugget 1991)
- El ecotono es la transición espacial entre dos o más comunidades distintas, y puede tener una extensión lineal considerable (Butzer 1985, Hugget 1991)
- En los modelos de selección de tipo K y selección de tipo r , los procesos denso-dependientes son los que ocurren en un contexto de alta densidad de organismos en su ambiente. En tales contextos las poblaciones tipo K son aquellas en las que la supervivencia depende en mayor grado de la competencia. Contrariamente, en las poblaciones de tipo r la densidad de individuos y la competencia no son factores importantes en la supervivencia, siendo los relevantes la inestabilidad y las fluctuaciones ambientales (Pianka 1982).
- Un deme es una Población Local con un hábitat particular en el área de distribución de una especie.
- La adaptividad potencial es una medida de la probabilidad de una entidad evolutiva de ser replicada, lógicamente independiente del proceso de selección (O’Brien y Holland 1992). En este marco una variante adaptativa es un estado del rasgo (estructuras biológicas, conductas, artefactos) que aumenta la probabilidad de supervivencia y reproducción al portador; y una variante No Adaptativa del mismo rasgo es aquella que le disminuye esta misma probabilidad.
- Los nunamiut explotan un área anual de 63.700 Km2 con una distancia promedio por movimiento residencial de 69,5 Km, en un bioma con PP de 73/g/m2/yr, y alta estacionalidad (Kelly 1983)
- Sin embargo puede argumentarse, como hace Madsen (1993), que las presas pequeñas mediante una táctica de caza con redes o trampas -u otras similares- pueden mejorar el rendimiento. No obstante, esto implica una estrategia de mucho menor beneficio frente a la que logra efectivamente consumir, con una mayor frecuencia, a los artiodáctilos. Un caso contrario podría ser la caza masiva.
- Esta línea de argumentación tiene sustento empírico y teórico cuando se coteja el comportamiento de la Biomasa de Herbívoros de los diversos biomas terrestres en relación a la Biomasa Primaria (McNaughton et al 1989, Pianka 1982), muy especialmente en los paisajes áridos como el de la Puna (Mc Naughton et al 1993, y particularmente para los camélidos (Rabinovich 1989).
- La etología de los camélidos es muy flexible en sus patrones de movilidad, por lo que debe ser muy sensitiva de la degradación de la cobertura vegetal. Frente a la disminución de las pasturas en el hábitat los camélidos aumentan la movilidad migratoria, para disminuirla en ambientes más ricos. En el caso de la vicuña esta flexibilidad le permite explotar una mayor rango ambientes áridos (ver Flannery et al 1989).
- El valor obtenido de esta manera corresponde a la Utilidad Energética por Tiempo de Procesamiento (Tpi) el cual es una fracción de la Tasa de Calorías por Tiempo de Manejo. Luego resulta de : Tpi = Ei / tpi .
- La recolección masiva de los huevos de suri puede llevar a que este recurso aumente levemente su posición en el ranking si se realiza el cálculo a partir de otras premisas u observaciones, respecto a los tiempos de procesamiento.
- El producto µ está compuesto por dos magnitudes de energía, por espacio: hectárea (Ht) y por tiempo: horas (Hrs) . Se unidad resulta : Kcal2 / Ht.Hrs. Es apropiado transformar este producto en un índice, normalizándolo al valor 106 , lo que permite trabajar sin unidades y con un número de dígitos manejables.
- El número de combinaciones posibles de series de recursos con sus tres estados de disponibilidad en el paisaje resulta de 39. Si bien esto está asumiendo un estatus equiprobable para cada variable cuando quizás muchas covarían, advierte la alta variabilidad esperada de ambientes de escala local.
- El gráfico no muestra esta situación pero puede demostrarse fácilmente con un cociente donde la media de µ sea el numerador y la superficie del rango el denominador, aumentando el número de ambientes de la serie. En teoría el máximo de ambientes incluidos se alcanza cuando los costos de la movilidad interceptan el rendimiento promedio de toda la serie.
Buscar en esta seccion :