 Noticias de Antropologia y Arqueología
Noticias de Antropologia y Arqueología
Colonización Humana del NOA y Variación en el Consumo de los Recursos:
La Ecología de los Cazadores Recolectores de la Puna Durante la Transición Pleistoceno- Holoceno. (2da parte)
Hernán J. Muscio
CONICET. Instituto de Ciencias Antropológicas (UBA) Sección Prehistoria. 25 de Mayo 217 4º Cap. Fed. Argentina. hmuscio@iname.com
Comentarios
Luis Alberto Borrero
Programa de Estudios Prehistóricos (PREP), CONICET
Bartolomé Mitre 1970 5º A Cap. Fed . Argentina
Con el trabajo de Hernán Muscio tenemos una de las primeras discusiones sobre maladaptaciones en la arqueología Argentina, tema que infiere a partir del tamaño de los recursos y de su rendimiento. Efectivamente estas son propiedades que típicamente hacen costosa una adaptación, y pueden llevar a que la proporción costos:beneficios sea demasiado alta.
La discusión sobre desvíos subóptimos en la dieta, sobre base arqueológica, es una contribución importante. La situación no sería llamativa, pues ocurre con casi todos los cazadores-recolectores estudiados por la etnografía desde la perspectiva de la ecología evolutiva. Esto confirma que en el mundo real no existen adaptaciones óptimas, y que, a lo sumo, hay diversos grados de acercamiento al óptimo. Podemos preguntarnos cuál es el margen de tolerancia que tiene una población. Este es uno de los grandes temas para investigación futura. Ante todo creo que aquí entran las "circunstancias", las formas de organización social, y otras propiedades difíciles de medir, pero que constituyen muchas veces las diferencias que se perciben entre poblaciones. Es claro que uno puede trabajar con un enfoque cuantitativo, y medir la subsistencia y otros temas, y aún disponer de lugar para evaluar decisiones llamadas "culturales". Tan solo que estas últimas nunca pueden constituir toda la explicación para un conjunto de conductas.
Pero quiero enfatizar cuan profundamente sociales pueden ser estos modelos, claro que de una manera diferente a la que planteaba la "arqueología social". No se trata aquí de asumir etapas en la organización social, inspirándonos en lo que ofrece la etnografía, sino en utilizar esos conocimientos para iniciar una búsqueda de lo desconocido, de estructuras de organización insospechadas. En esto está pensando Beaton al especular, refiriéndose a los tempranos colonizadores de un continente, que sus "marriage and residence rules might be practically unreasonable, if not altogether demographically dangerous" (Beaton 1990: 34). Potencialmente, la duración de una generación puede variar mucho, de acuerdo con los principios de parentesco que estén funcionando. Es claro que distintas formas "poco óptimas" de relacionarse con el ambiente (incluyendo los vecinos) pudieron generar, o requerir, organizaciones sociales especiales. Por ejemplo, está muy poco explorada la existencia de sistemas endogámicos, cuya duración en el tiempo puede ser de pocas generaciones, y algunas de cuyas implicaciones arqueológicas son describibles. Puede pensarse en una demografía sumamente baja como en una condición creadora de tales sistemas y puede pensarse en altas tasas de endogamia como resultado. Recientemente Sanguinetti de Bórmida y otros (1998: 442) presentaron restos osteológicos humanos de dos individuos para la cuenca del río Limay, para los que se ha sugerido que: "La similitud morfológica natural de los cráneos hace suponer la existencia de un estrecho parentesco entre ambos individuos o una elevada endogamia en el grupo de pertenencia".
La inestabilidad de los ambientes del Pleistoceno final, que es cuando están comenzando a ser colonizados muchos sectores de Sud América, es un rasgo importante en esta discusión (Clapperton 1993). Su importancia está subrayada porque los grupos que pudieron estar participando de la exploración de la Puna o de otros sectores del espacio hacia fines del Pleistoceno debían ser particularmente débiles, dado que se daban muchas condiciones para que se cortaran los vínculos con las poblaciones de las que se estaban desprendiendo. Bajo un modelo de lento crecimiento demográfico, en la línea de Beaton (1990), cualquier desbalance con los recursos significaba la posibilidad de extinción. Esto ocurriría aunque los grupos estuvieran bien adaptados; si consideramos que Muscio nos dio buenos elementos para sugerir que no siempre lo estaban, entonces la situación empeora. En términos ecológicos en realidad es esperable que haya inicialmente un éxito inmediato (Edwards y O’Connell 1995: 776), pero la realidad es que en varios tipos de ambientes, usualmente tratados como "marginales", la situación se puede acercar a la sugerida por Beaton. Entonces, la falta de un boom demográfico es lo esperable, y esto sería lo que precisamente estaría ocurriendo –en una escala regional. Allí reside la posibilidad de una maladaptación. Pero aún en el caso de un éxito demográfico inicial, usualmente a expensas de los recursos de más alto ranking, es esperable un decrecimiento poblacional producido por el costo del posterior acceso a recursos más costosos. El desafío para los arqueólogos es distinguir entre ambos tipos de situaciones.
La discusión acerca de continuidad o discontinuidad en el uso de ciertos sectores del espacio regional (Yacobaccio 1998), ya en el Holoceno medio, forma parte integral de los temas que pueden abordarse con este enfoque, dependiendo de la creación de una adecuada base de datos paleoecológicos.
José Luis Lanata
Dep. de Antropología, UBA y
Programa de Estudios Prehistóricos (PREP), CONICET
jllanata@filo.uba.ar
Ha diferencia de lo pensado hasta hace pocos años atrás, hoy sabemos que la transición Pleistoceno-Holoceno - llegando incluso hasta el Holoceno medio- llevó consigo cambios importantes en los ambientes Sudamericanos. Por otra parte, nuestro continente se caracteriza por haber tenido -y tener- una importante diversidad de ecosistemas, lo que sumado a lo dicho anteriormente, nos hace pensar que la dispersión y colonización humana tiene que haber tenido un tempo y una forma particular en cada región, incluso con pulsos diferenciales en ellas.
El trabajo de Muscio presenta una serie de aspectos relevantes relacionados con este tema, desarrollado en este caso para la Puna. Por un lado, muestra que la maladaptación fue uno más de los procesos que tuvieron lugar en la historia de las poblaciones de cazadores-recolectores sudamericanos -así como de otros tipos de subsistencia. Esto sin duda nos habilita aún más a pensar que la extinción -local y/o regional- de poblaciones humanas fue un resultado no tan infrecuente como muchos piensan -ver el comentario de Borrero.
El otro punto que Muscio deja vislumbrar es como el espacio, considerándolo desde un punto de vista ecológico y biogeográfico, actúa diferencialmente en distintos procesos evolutivos, como el mencionado de dispersión y otros -i.e. transmisión cultural, variación demográfica. En la arqueología, tan solo el caso de las barreras ha sido considerado, viéndolos como sectores que alteraban y/o modificaban -al menos- la movilidad de las poblaciones. Sin embargo, se ha olvidado que la relación espacio-tecnología hace que esas barreras dejen de ser tales, actuando como filtros, corredores y/o sweeptakes (Lanata y Neff 1998). Barreras acuáticas -ríos, canales, estrechos e incluso los océanos-, terrestres -cordones montañosos, depresiones-, ambientales -selvas, desiertos-, entre otras, han sido superadas en diferentes momentos y circunstancias, por nuestra especie ya sea a lo largo del proceso de dispersión global como en los diferentes casos locales y regionales. De esta forma, las preguntas que debemos formularnos en el caso que presenta Muscio, como en otros semejantes -i.e. las Rocallosas, Los Andes- son por cuánto tiempo la Puna fue una barrera y bajo que condiciones -ambientales, tecnológicas, etc.- fue superada. Esto puede comenzar a trabajarse con el tipo de información presentada en el trabajo. Y de allí en más surge otro tema, que rol jugó posteriormente para las poblaciones humanas. Fue sencillamente un espacio más de los explotados, y/o pudo actuar de alguna de las formas mencionadas? Esto puede comenzar a analizarse también en la forma que Muscio lo plantea, complementando los enfoques seleccionistas y ecológico evolutivos en arqueología.
A partir de conocer esta variabilidad en el rol que el espacio comienza tener en diferentes procesos evolutivos, comenzamos a ver que la dinámica de dispersión y colonización de nuestra especie parece haber sido mucho más versátil de lo que pensábamos. Y viendo estas posibilidades, comenzamos a replantearnos los modelos que usamos para su interpretación. Fundamentalmente, los que empleamos -i.e. flujo lento y/o rápido- son los más simples de la dinámica de fluidos y no parecen ser los adecuados para el de dispersión humana (Foley com. pers.). Nuevos modelos que den cuenta del tempo y modo de la dispersión así como de la diversidad ecológica de los diferentes espacios parece ser el camino que debemos explorar. De allí surgen temas de demografía, conflictos, competencia y/o intercambio de recursos, transmisión de rasgos, que esperan ser atacados. La propuesta de Muscio además de mostrarnos la necesidad de replantearnos algunos modelos de uso el espacio en la Puna, y de testerar otros, plantea la necesidad de indagar sobre estos temas.
Anexo
Abundancia, Utilidad y Variaciones en el Consumo de los Recursos: Hipótesis de Movilidad y Amplitud de La Dieta Prehistórica
Se puede argumentar que la escasez y la irregularidad en la oferta de los recursos debieron haber sido los principales estresores de la subsistencia de las poblaciones humanas prehistóricas en la Puna. Estas dos propiedades de la estructura de los recursos requieren diferentes tratamiento analítico (ver Nelson 1996). Bajo criterios de "optimización" la adaptividad del comportamiento en biomas de escasez de recursos -tundras y desiertos en general- se puede analizar mediante modelos de maximización del beneficio. El modelo de Amplitud de Dieta es adecuado para esto. Con respecto a la irregularidad, considerando su dimensión espacial y temporal, son apropiados los modelos de uso heterogéneo del espacio y los de manejo de fluctuaciones en contextos de riesgo (Cashdan 1990, Escola 1996, Lanata y Borrero 1994, Smith 1988).
En un trabajo anterior (Muscio 1998a) se presentó un ranking del rendimiento de los recursos alimenticios de la Puna, a partir de la utilidad post-encuentro de cada uno de ellos. La tabla 1 reproduce esa información. (11)
|
RECURSOS |
PESO |
UTILIDAD NETA Kg |
TIEMPO DE |
TP |
NORMAL |
RANK |
| Taruca Guanaco Vicuña Suri Tubérculos Huevos de Suri Vizcacha Frutos Aves Hierbas Semillas |
50 70 45 20 1 0,60 1,6 1 1,4 1 1 |
29 42 27 13 1 0,6 1,2 1 0,91 1 1 |
0,02 1 0,02 1,40 0,02 0,90 0,05 1 0,08 0,08 0,08 0,08 0,15 0,25 0,08 0,08 0,18 0,25 0,08 0,08 5 5 |
36.250 32.400 32.400 24.700 16.625 11.850 8.108 8.000 6.916 1.750 323 |
1 0,89 0,89 0,68 0,45 0,32 0,22 0,22 0,19 0,05 0,01 |
1 2 2 3 4 5 6 6 7 8 9 |
Tabla 1: Ranking del rendimiento post-encuentro de los recursos de la Puna, normalizado a partir del máximo valor = taruca (Muscio 1998a).
El Modelo de Amplitud de Dieta predice que la inclusión de la biomasa de bajo ranking depende de la abundancia en el paisaje de la biomasa de alto ranking. Si los recursos rankeados alto son muy frecuentes en el paisaje los individuos que se aproximan a soluciones óptimas ignoran los restantes. Paralelamente, en el caso de los recursos que pueden ser recolectados en masa (semillas, peces, insectos, animales pequeños en general), la utilidad post encuentro puede ser una función de la densidad de estas presas en el paisaje (ver Madsen y Schmitt 1998). En estos alimentos la unidad de comparación no es la presa individual, sino un volumen de captura masiva. De tal manera, en situaciones particulares, estos recursos de recolección masiva pueden transformarse en presas de alto ranking. Los huevos de suri son un buen ejemplo de esta clase de recursos, dado que están disponibles en un número de 8 a 25 por nido, localizados en pastizales bien predecibles durante la primavera (Muscio 1998a). (12)
Pueden considerarse dos variables arqueológicamente discutibles de la abundancia de los recursos: biomasa y distribución (Yacobaccio 1994). Estas variables afectan la frecuencia de encuentro de los recursos, controlando los costos de búsqueda (en energía y tiempo) de los alimentos. El Modelo de Amplitud de la Dieta predice que la secuencia de inclusión de los recursos se interrumpe cuando los costos de búsqueda igualan a la utilidad neta. Esto gráficamente se observa en la intersección de ambas curvas ( D t ) del modelo básico (ver Smith 1983). Los datos que en este trabajo se construyen no tienen esa resolución, especialmente en los costos de búsqueda como una función del tiempo. Interesa evaluar la mejor alternativa para la subsistencia humana en función de la utilidad post- encuentro de los recursos (TP) y de la oferta ambiental de cada uno de ellos. Se busca establecer la secuencia óptima de inclusión de los recursos en la dieta pero sin fijar su amplitud máxima, donde se interrumpe la entrada de los recursos de bajo ranking. Se sigue esta metodología para derivar hipótesis de estrategias óptimas, subóptimas y no adaptativas en el comportamiento de subsistencia. Se parte estableciendo la relación µ entre la disponibilidad de los recursos (D) y la utilidad económica de ellos (TP) un producto, dado por :
µ = Tp (Di )
Este producto indica la utilidad neta de la energía de la que se dispone en el ambiente y discrimina aquellos recursos que ofrecen alta disponibilidad y alto rendimiento, de los que ofrecen baja disponibilidad y bajo rendimiento, resultando un estimador de la utilidad de la biomasa ambiental. El término Di es la disponibilidad por unidad de área de un recurso i . En este planteo es conveniente uniformar las medidas de disponibilidad ambiental en unidades comparables: Kilocalorías. De aquí Di resulta una medida de la biomasa dada en su valor bruto de energía (el producto entre la cantidad de recurso i expresado en Kg/Ht y el valor energético de esa biomasa según las tablas), su unidad entonces resulta Kilocalorías/Ht (13). Asumiendo una relación lineal (en el sentido matemático del término) entre la frecuencia de encuentro y el tiempo de búsqueda, los valores más altos de µ indican la mejor opción en el paisaje. Dado que son los de mayor utilidad con mayor frecuencia de encuentro resultan los de mayor rendimiento por tiempo de búsqueda, ordenados de mayor a menor se obtiene la serie de la secuencia óptima con la cual deben entrar en la dieta los recursos. Se deduce entonces que µ es un estimador de la adaptividad del consumo de los recursos en ambientes específicos de escala local.
Heterogeneidad Espacial y Ambientes Locales :En la escala local la heterogeneidad espacial de la distribución de los recursos de la Puna se expresa como agregaciones en sectores discretos del paisaje, tales como las vegas, las quebradas protegidas o las lagunas. Para captar esta heterogeneidad, el método que se emplea es la modelización de los ambientes locales mediante cantidades ideales de biomasa de cada recurso. A tal fin se utilizan los valores extremos y medios de la variación de la biomasa de cada recursos en una escala regional, tomados de otros trabajos (Muscio 1996,1998), para calcular sobre estos el índice µ. El resultado es una estimación de la utilidad de la biomasa, en situaciones de oferta mínima, media y alta de cada recurso. La tabla 2 presenta la variables consideradas y los valores µ calculados.
| RECURSOS |
DISPONIBILIDAD |
µ |
µ |
µ |
|
| media | ± ( %) | ||||
| Guanaco Vicuña Taruca Vizcacha Suri Tubérculos Hierbas Frutos Semillas |
234,36 150,66 256,25 463,36 160 65,17 12,18 105,10 96 |
96 60 83 89 68 80 80 80 80 |
0,30 |
7,60 |
14,88 |
Tabla 2
De la tabla 2 se extraen los datos necesarios para modelar los ambientes locales. Esto se realiza tomando un único valor µ para cada recurso y armando una serie completa (14). Este procedimiento permite analizar como las alternativas de optimización son una función de la abundancia relativa de los recursos en el paisaje. En este trabajo se considera un número limitado de series de ambientes locales para estudiar situaciones específicas, comenzando con la movilidad y la escala espacial de hábitat.
Movilidad y Escala del Hábitat de los Cazadores Recolectores: Las ocupaciones de los cazadores recolectores en la Puna se emplazan en ambientes locales de concentración relativa de nutrientes. Como demostró Yacobaccio (1991,1994) estos loci pueden definirse como Zonas de Concentración de Nutrientes. Interesa subrayar que las ZCN permiten estudiar el rango de acción mediante modelos de uso óptimo de ambientes heterogéneos, ya que se pueden tratar como parcelas óptimas.
Existe consenso en una estrategia de movilidad muy alta de los cazadores recolectores de la Puna. Esta hipótesis se puede sostener desde la Teoría de Uso Óptimo de Ambientes de Mosaico (Mac Arthur y Pianka 1966). En este modelo el rol funcional de la movilidad es promediar de manera óptima la varianza entre ambientes distintos (Cashdan 1990). La selección actúa favoreciendo los movimientos en el paisaje que rinden, promediadamente, el mayor beneficio. De la misma manera que en el Modelo de la Amplitud de la Dieta, la serie de los ambientes explotados (parcelas óptimas) incluye secuencialmente nuevos ambientes hasta que el costo de la movilidad no supera la utilidad promedio. La ventaja descansa en la optimización del rendimiento de los recursos capturados mediante la movilidad entre distintos ambientes locales del espacio regional, en función de los costos que esta actividad insume. Dado que en la Puna la varianza de la oferta de los recursos en el espacio es alta, por una distribución muy heterogénea en distintas escalas espaciales el umbral óptimo determina un rango extenso y una alta movilidad, con la explotación de varios ambientes locales (Muscio 1998b).
La figura 1 grafica una situación donde 3 ambientes -ZCN- son explotados en el rango anual: A,B,C. Cada uno de los ambientes tiene un promedio de utilidad específico, dado por la densidad de sus recursos medida con µ . En el ejemplo del gráfico los tres tienen promedios similares en orden de magnitud, pero de distinta composición. El ambiente A es medianamente rico en cérvidos, camélidos y tubérculos. B tiene una buena disponibilidad de suri, vicuña y herbívoros de bajo ranking. C tiene una muy alta riqueza de cérvidos, pero dentro de una baja densidad de biomasa de alto ranking. Cada uno de estos ambientes presenta distintos espacios de caza y recolección de recursos.
En el gráfico la línea de mayor intensidad representa el promedio de utilidad del rango ABC. En una escala de ± 2.500 Km2 el rango ABC es un buen modelo para rangos de Puna y Prepuna, donde C puede ser un ambiente local en la Prepuna y las Quebradas de acceso, mientras A y B pueden localizarse en el valle de San Antonio de los Cobre en la Puna de Salta (ver Muscio 1996). Cada nuevo ambiente que se agrega altera el rendimiento promedio de toda la serie (15). A partir de este modelo se observa que la amplitud del rango va a estar controlada por la densidad y por la diversidad local de los recursos. Dejando constante el tamaño de la población, los ambientes más ricos y diversos en recursos requieren una menor movilidad en un hábitat de menor escala espacial, por la inclusión de un menor número de ambientes locales para la realización de la dieta.
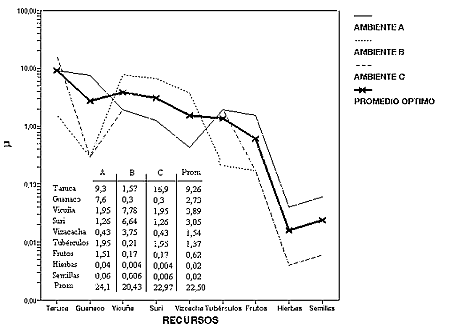
Figura 1: Rendimiento promedio de los recursos en un rango regional de caza y recolección hipotético, definido por tres ambientes locales .
Focalizando en la variación de las frecuencias de encuentro de los recursos -gobernada por la biomasa- respecto de la selectividad humana de presas se analizan tres clases posibles de conductas: A) Comportamientos de Alto Beneficio B) de utilidad Subóptima, C) de Bajo Beneficio, No Adaptativa. La primera prioriza el rendimiento neto de los recursos para su inclusión en la dieta, la segunda es un desvío de la optimización neta que prioriza la disponibilidad ambiental por sobre el rendimiento post-encuentro de los recursos. La tercera explota una base de recursos no óptima y promueve la maladaptación.
Comportamientos de Alto Beneficio: La figura 2 grafica la nube de puntos de una disponibilidad media, máxima y mínima de biomasa útil en una dieta humana en la Puna. El gráfico toma los valores de la tabla 2 , expresando el eje y en una escala logarítmica para no perder visualmente los valores mínimos de las tres series de puntos. La serie A representa los valores µ de los recursos cuando la disponibilidad es la media regional para todos los componentes de la serie. En esta situación la conducta óptima, que predice el Modelo de Amplitud de la Dieta, comienza incluyendo taruca -el recurso de mayor beneficio- continúa con guanaco, vicuña, suri y finalmente los recursos pequeños, vizcacha, tubérculos, semillas, frutas y hierbas. Dado que la serie está calculada a partir de la oferta promedio de los recursos en la Puna, constituye una hipótesis de la inclusión óptima de los recursos en una escala regional. De tal manera los comportamientos de Alto Beneficio van a incluir, en los promedios de largo plazo, principalmente a los herbívoros de mayor tamaño , y con mucha menor frecuencia presas pequeña y biomasa vegetal. A continuación se analizan series de hábitats de escala local, para poblaciones de cazadores recolectores.
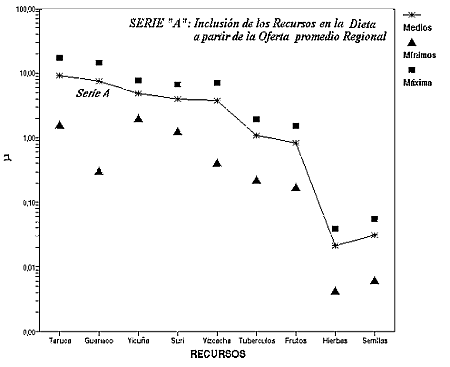
Figura 2: Rendimiento de los recursos en función de la oferta promedio regional en la Puna
De la nube de puntos de la fig. 2 se toma una serie para modelar un ambiente de escala local con una alta frecuencia de biomasa de alto ranking, simulando una situación de optimización del rendimiento.
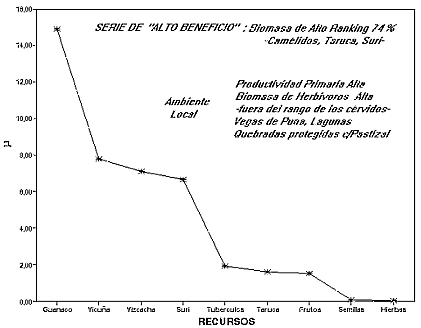
Figura 3: Serie de Alto Beneficio, a partir del consumo de los recursos de un ambiente local
La figura 3 presenta este caso, con una serie local coherente con la curva de optimización regional. En este ambiente local la oferta de biomasa primaria es alta, llevando a una biomasa de herbívoros también alta, salvo para taruca cuya biomasa es mínima exigiendo entonces una mayor inversión en tiempo de búsqueda. Esta situación modela un ambiente de Puna con buena oferta de PP y humedad relativa en lugares discretos de escala local -que podrían tener hábitats en la prepuna-. En esta situación los camélidos son la mejor opción seguidos por los roedores y el suri en niveles similares. Se observa que esta serie deja a taruca en el mismo nivel en el que están los recursos vegetales.
El ejemplo muestra que el paisaje permite la optimización a partir del rendimiento neto de los recursos con una alta frecuencia de consumo de los recursos de mayor utilidad post-encuentro. Suponiendo que esta serie representara en valores absolutos la biomasa consumida en la dieta se observaría que el 74 % estaría compuesta por recursos de alto Alto Ranking, aunque la inclusión de los roedores sea abundante. Esta situación puede explicar las ocupaciones del Holoceno Temprano de Huachichocana (E 3) en Jujuy y de Quebrada Seca (QS3) en Catamarca.
Comportamientos Subóptimos: Una estrategia subóptima implica la inclusión muy frecuente de recursos de mediano ranking en la dieta por la escasez de los recursos de más alto ranking. La estrategia subóptima está controlada por la disponibilidad local de los recursos y luego por el rendimiento post-encuentro de ellos (ver Bettinger 1993). Un comportamiento subóptimo no se aleja demasiado del óptimo, dado que depende de los costos de oportunidad. Cuando el costo de oportunidad de un recurso es alto -supera el beneficio o se acerca a él- su captura es una mala alternativa. El beneficio neto ganado si se toma el recurso de alta disponibilidad pero subóptimo en el ranking y se rechazan los otros, depende de la magnitud de la diferencia entre la utilidad que se toma y la que se rechaza. Cuando la diferencia entre la utilidad neta de los recursos es muy grande, lo que se pierde por tomar el recurso alternativo (el más frecuente) es mucho; dado esto la estrategia más beneficiosa resulta ignorar la disponibilidad actual de bajo rendimiento y seguir buscando el recurso de mayor tasa de utilidad post-encuentro si existen expectativas favorables de encontrarlos. Es por esta razón que los comportamientos subóptimos no se alejan demasiado de los óptimos y manejan recursos de mediano ranking.
La figura 4 grafica una serie donde la disponibilidad local de biomasa de herbívoros es mediano-baja, pero con una biomasa primaria que ofrece hábitats para las presas de tamaño mediano - suri y vicuñas -. Esto modela una situación donde guanaco y vicuña no superponen totalmente su rango, mientras suri sí lo hace hasta por lo menos 4.000 m/nm. En la serie suri es el recurso de mayor beneficio por su disponibilidad local, pero subóptimo en el ranking post-encuentro. Volviendo a la Tabla 1 NORMAL, se observa que la distancia de suri al recurso de mayor ranking -taruca- es de 0,32 (1 - 0,68). Luego secuencialmente entran: vicuña, con una distancia de 0,11 y tubérculos con una distancia de 0,53. Este último recurso marca el límite de lo subóptimo. Obsérvese que en esta serie la biomasa de alto ranking representaría el 80 % del total, si fuera consumida de manera proporcional a la curva. Esta serie local resulta entonces una desviación subóptima de la serie de Alto Beneficio. La inclusión de componentes subóptimos en la dieta es compatible con una conducta diversificadora en una estrategia que amortigua de manera oportunista la fluctuación en la disponibilidad de las presas de caza. Para los cazadores recolectores de la Puna esto implica que una serie óptima puede incluir desvíos subóptimos, funcionales en contextos de riesgo, complementando la estrategia de movilidad y uso del espacio en un rango de acción amplio.
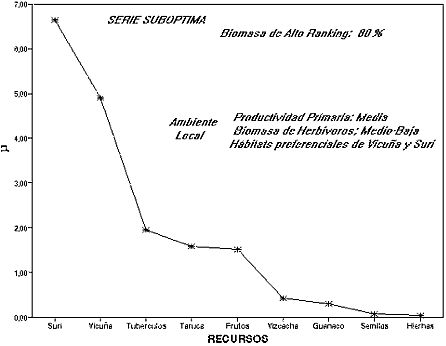
Figura 4: Serie subóptima a partir de la inclusión de recursos de mediano rendimiento y buena disponibilidad en un ambiente local.
En una escala global la etnografía demuestra que los comportamientos oportunistas evitativos del riesgo, o risk averse, son una parte fundamental de la estrategia de subsistencia, en la mayoría de las poblaciones de cazadores recolectores de los distintos biomas del planeta. Por regla las fluctuaciones en la caza de presas de alto rendimiento, generalmente mamíferos terrestres mediano/grandes, se amortiguan mediante la inclusión de semillas y recursos más pequeños de mayor predecibilidad y accesibilidad (ver Kelly 1983). Este sería el patrón observado en la mayoría de las sociedades etnográficas, incluyendo los !Kung del Kalahari (Cashdan 1990, Wiessner 1977) y los Aché de las selvas de Paraguay. De tal manera, la estrategia de caza y recolección parece encontrar un potencial adaptativo mayor en la flexibilidad del comportamiento oportunista de consumo de recursos óptimos y subóptimos, que en las especialización en una serie estrecha de presas de caza. Esta flexibilidad es también importante en los procesos de dispersión hacia nuevos hábitats, debido a que posibilita la explotación de una mayor diversidad de ambientes. Ciertamente, un comportamiento demasiado especializado restringe la realización del nicho a una geografía con propiedades ecológicas más homogéneas, similares a las de origen. Sobre esta base se postula que la estrategia de dispersión humana en el continente americano durante la Transición Pleistoceno Holoceno, se habría caracterizado por un uso oportunista de los recursos, a medida que la expansión iba incorporando los nuevos y diversos ambientes al rango colonizado (ver Kelly y Tood 1990).
Comportamientos de Bajo Beneficio y No Adaptativos: Winterhalder (1993) matemáticamente demostró, mediante la simulación dinámica del comportamiento demográfico de grupos de cazadores recolectores presionando su base de recursos, que bajo un contexto de disminución de la disponibilidad de los nutrientes, la estrategia de mayor ventaja adaptativa es la disminución del tiempo dedicado a la predación y no la inversa. Estos contextos locales favorecen una intensificación de la energía producida y consumida por unidad de área en el hábitat, que se logra disminuyendo la energía y el tiempo invertida en la subsistencia. Esto se logra disminuyendo la búsqueda de los recursos de alto rendimiento y ampliando la base de los recursos explotados, privilegiando a los de mayor abundancia neta por sobre los de alto beneficio post-encuentro. Sin embargo, en el largo plazo, el aumento de la amplitud de la dieta y de la frecuencia de inclusión de recursos de bajo rendimiento puede conducir a situaciones No Adaptativas, con implicancias tanto en el perfil sanitario de la población como en sus patrones demográficos:
«...all else being equal, foragers who maximize nutrient returns rates enjoy greater reproductive succes than those who do not. Increses in diet dreadth result from reduced foraging return rates an so lead to declines in population growth rates» (Hawkes and O’Connel 1992:62).
La figura 5 grafica la serie óptima para un ambiente de escala local de productividad primaria mediana baja, pero con alta disponibilidad de los vegetales para una dieta humana. Este ambiente presenta una baja densidad de herbívoros de alto ranking y una alta disponibilidad de animales de bajo rendimiento con hábitats bien localizables en el paisaje. Este es el caso de los chinchíllidos, con hábitats en los pastizales de baja PP.
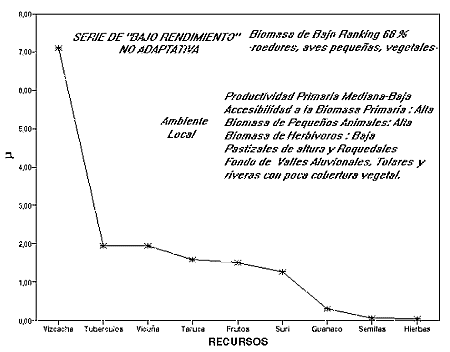
Figura 5: Serie No Adaptativa de consumo de recursos,
a partir de la escasez local de los alimentos de alto rendimiento
La baja densidad de carne de alto ranking impone una ampliación de la dieta en la cual vizcacha es el recurso con mayor frecuencia de entrada, seguido por los tubérculos y vicuña -en el mismo nivel- y el resto de los recursos. Esta estrategia implica la inclusión de presas de bajos costos de búsqueda relativos, pero de bajo rendimiento post-encuentro. En esta serie los recursos de alto ranking constituyen solo el 32 %, contra los de bajo ranking -muy alejados del óptimo- presentes en el 68 % del total. Esta serie puede explicar el registro de Inca Cueva 4 C2, y probablemente el de las primeras ocupaciones del Alero de las Circunferencias.
En los individuos la consecuencia probable de las variantes no adaptativas es la maladaptación que denota la expresión biológica en el organismo del desajuste a las restricciones ambientales (ver la discusión de Holland y O’Brien 1997). En la ontogenia la maladaptación afecta a la esperanza de vida y al potencial reproductivo del individuo y, en cascada, a la estructura de la población (ver Jackes et al 1997).
En una población de escala regional, los rasgos no adaptativos y las maladaptaciones en general promueven procesos de extinción o extirpación (extinción local) de fracciones de población -demes o grupos- en el largo plazo. Un régimen de selección contra los comportamientos no eficientes en la captura manejo y consumo de los recursos, puede llevar a la extirpación de poblaciones humanas de ambientes colonizados o en proceso de colonización de ambientes marginales (ver Mandryk 1993). En estas situaciones pueden estar presentes diversos factores tales como las limitaciones en el hábitat por la baja productividad del entorno, pulsos impredecibles de escasez de recursos (Cashdan 1990), superposición de nichos y competencia (Bettinger y Baumhoff 1982), o la modificación de la ecología local por la sobreexplotación de los recursos desde un lugar central (Broughton 1997, Janetski 1997) .
Bibliografía Citada:
Adams, J., M, and H. Faure
1997 Preliminary Vegetation Maps of the World since the Last Glacial Maximun: and Aid to Archaeological Understanding. Journal of Archaeological Science 24:623-647.
Aguerre, A., A. Fernández Distel, y C. Aschero
1975 Comentarios sobre nuevas fechas en la cronología arqueológica precerámica de la Provincia de Jujuy. Relaciones de la Sociedad Argentina de Antropología (N.S.) IX:211-214. Buenos Aires.
Aschero, Carlos, A
1984 El Sitio ICC-4: Un Asentamiento Precerámico en la Quebrada de Inca Cueva (Jujuy, Argentina). Estudios Atacameños 7:62-72.
1994 "Reflexiones Desde el Arcaico" -Actas y Memorias del XI Congreso Nacional de Arqueología Argentina, Revista del Museo de Historia Natural de San Rafael - (Mendoza). Tomo XIII (1/4) San Rafael pp13.
Avery, D., M
1995 Physical Environment and Site Choice in South Africa. Journal of Archaeological Science 22:343-353.
Beaton, J.M., 1990. The Importance of Past Populations for Prehistory. Hunter-Gatherers Demography. Past and Present (Eds. B. Meehan and N. White), pp. 23-40, University of Sydney
Bettinger, Robert, L
1993 Doing Great Basin Archaeology Recently: Coping with Variability. Journal of Antropological Research 1 (1) :43-66
Bettinger, Robert, L, and Martin Baumhoff A
1982 The Numic Spread: Great Basin Cultures in Competition. American Antiquity 47(3).
Bettinger, Robert, L, Ripan Malhi, and Helen McCarthy
1997 Central Place Models of Acorn and Mussel Processing. Journal of Archaeological Science 24:887-899.
Bianchi, A., R, y C. Yañez E
1992 Las Precipitaciones en el Noroeste Argentino. 1 ed. INTA, Salta.
Binford, Lewis, R
1994 Human Actors and Their Role in the Evolutionary Play, MS.
Bonzani, Renée, M
1997 Plant Diversity in the Archaeological Record: A Means Toward Defining Hunter-Gatherer Mobility Strategies. Journal of Archaeological Science 24:1129-1139.
Borrero, Luis, Alberto.
1988 Problemas en la Definición de Sistemas Adaptativos. Arqueología en las Américas. pp. 248-256. Bogotá.
1990 Evolución Cultural Divergente En La Patagonia Austral. Anales del Instituto de la Patagonia 19:133-140.
1993 Artefactos y Evolución. Palimpsesto 3:15-32, Buenos Aires.
1995 Arqueología de la Patagonia. Palimpsesto, Revista de Arqueología 4:9-66.
1997 La Extinción de la Megafauna en la Patagonia. Anales del Instituto de la Patagonia 25:89-102, Ser. Cs. Hs.
Borrero, Luis, Alberto., y Nora Franco Viviana.
1997 Early Patagonian Hunter-Gatherers: Subsistence and Technology. Journal of Anthropological Research 53:219-239.
Broughton, Jack, M
1997 Widening Diet Breadth, Declining Foraging Efficiency, and Prehistori Harvest Pressure: icthyofaunal evidence from the Emerville Shellmound, California. Antiquity 71:845-862.
Broughton, Jack, M, and Donald Grayson K
1993 Diet Breadth, Adaptive Change, and the White Mountains Faunas. Journal of Archaeological Science 20:331-336.
Butzer, K., R
1985 Archaeology as Human Ecology. Cambridge University Press, Cambridge.
Cashdan, Elizabeth
1990 Spatial Organization, and Habitat Use. In Evolutionary Ecology and Human Behaviour, pp. 237-267. Aldine de Gruyere, New York.
Cigliano, Eduardo, Mario.
1965 Nuevos Sitios Precerámicos en la Puna Argentina. En Anales de la Comisión de Investigación Científica Vol. VI La Plata.
1966 Panorama General de las Industrias Precerámicas del Noroeste Argentino. En Congreso Internacional de Americanistas - 37, vol. 3. Actas y Memorias., Buenos Aires.
Clapperton, C.
1993 Quaternary Geology and Geomorphology of South America. Amsterdan: Elsevier Science Publishers
Dunnel, Robert, C and William Dancey, S. 1983 The Siteless Survey: A Regional Scale Data Collection Strategy. Advances in Archaeological Method and Theory, Vol 6 Academic Press, NY
Edwards, D. y J.F. O’Connell
1995. Broad Spectrum Diets in Arid Australia. Transitions. Pleistocene to Holocene in Australia and Papua New Guinea (Eds. J. Allen and J.F. O’Connell), pp. 769-783, Antiquity 69, Special Number 265
Eldredge, Niles
1989 Macroevolutionary Dinamics, species,niches, & adaptive peaks. 1st ed. McGrow-Hill, New York.
Elkin, Dolores, C
1994 Subsistencia en la Quebrada de Pintoscayoc (Jujuy) en el Holoceno Temprano, Actas y Memorias del XI Congreso Nacional de Arqueología Argentina -San Rafael- Mendoza.
Escola, Patricia, S
1996 Riesgo e Incertidumbre en Economías Agro-pastoriles: Consideraciones Teórico-Metodológicas. Arqueología 6:9-23.
Fernandez, Jorge, Vera Markgraf, Hector Panarello O, Miguel Albero, Fernando Angiolini E, y Susana Valencio
1991 Late Pleistocene/Early Holocene Envioronments and Climates, Fauna, and Human Occupation in the Argentine Altiplano. Geoarchaeology: An International Journal 6:251-272.
Fernández Distel, Alicia
1974 Excavaciones arqueológicas en las cuevas de Huachichocana, Dpto. de Tumbaya, Provincia de Jujuy. Relaciones de la Sociedad Argentina de Antropología (N.S.) VII:101-127.
1978 Nuevos Hallazgos Precerámicos en la región de las Salinas Grandes, Puna de Jujuy, Argentina. Revista Instituto de Antropología VI(Universidad Nacional de Córdoba).
1980 Los Fechados Radiocarbónicos en la Arqueología de la Provincia de Jujuy. Fechas Radiocarbónicas de la Cueva III de Huachichocana, Tiuiyaco e Inca Cueva, Argentina Radiocarbono en Arqueología.
1986 Las Cuevas de Huachichocana, su Posición dentro del Precerámico con Agricultura Incipiente del Noroeste Argentino. Beitrage sur Allgemeinen und Vergleichenden Archaeologie 8:353-430.
Flannery, Kent, V, Joyce Marcus, and Robert Reynolds G
1989 The Flocks of the Wamani. Academic Press, .
Foley, Robert, A
1985 Optimality Theory In Anthropology. Man 20:222-242.
1989 The Ecological Conditions of Speciation: a Comparative Aproach to the Origins of Anatomically-Modern Humans. In The human revolution:behavioural and biological perspectives on the origins of modern humans, edited by P. Mellars A, and C. Stringer B, pp. 298-320. Edimburgh University Press, Edimburgh.
1996 Causes and Consequences in Human Evolution. Journal of the Royal Anthropological Institute 1:67-86.
Gamble, Clive
1993 Ancestors and Agendas. In Archaeological Theory: Who Sets the Agenda?, edited by Norman Yoffee, and Andrew Sherratt, pp. 39-52. Cambridge University Press, Cambridge.
1994 Timewalkers. The Prehistory of Global Colonisation. Alan Sutton. Stroud.
Gnecco, Cristobal
1995 Paleoambientes, Modelos Individualistas y Modelos Colectivos en el Norte de Suramérica. Gaceta Arqueológica Andina 24:5-11.
Graf, Kurt
1992 Pollendiagramme Aus Den Anden, vol. 34. Physische Geographie 34. Universität Zürich, Zürich.
Hammel, E., A, and N. Howell
1987 Research in Population and Culture: An Evolutionary Framework. Current Anthropology 28:141-159.
Hawkes, K., and J. O'Connell
1992 On Optimal Foraging Models and Subsistence Transitions. Current Anthropology 33(1):63-66.
Hernandez Llosas, María Isabel, Hubert Alem, María Falchi Pía., Mariel Gavilán, Hercilia Knauth, Juan Leoni Bautista., Pablo Masci, Graciela Scarafia, Diana Tamburini, y Damian Vainstub
1994 Proyecto Arqueológico Pintoscayoc. Departamento de Humahuaca, Provincia de Jujuy, Actas y Memorias del XI Congreso Nacional de Arqueología Argentina. Tomo XIV. San Rafael - Mendoza.
Holland, Thomas, D, and Michael O'Brien J
1997 Parasites, Porotic Hiperostosis, and The Implications of Changing Perspectives. American Antiquity 62(2):183-193.
Huggett
1991 Climate, Earth Processes and Earth History, vol. 1. Springer-Verlag, Berlin.
Ingold, Tim 1993 The Temporality of the Landscape. World Archaeology 25(2):152-174.
Jackes, Mary, David Lubell, and Christopher Meiklejohn
1997 Healthy but Mortal: Human Biology and the First Farmers of Western Europe. Antiquity 71:639-658.
Janetski, Joel, C
1997 Fremont Hunting and Resource Intensification in the Eastern Great Basin. Journal of Archaeological Science 24:1075-1088.
Kelly, Robert
1983 Hunther-Gatherer Mobility Strategies. Journal of Anthropology 39:207-306.
Kelly, Robert, L, and Lawrence Tood C
1988 Coming Into The Country:Early Paleoindian Hunting and Mobility. American Antiquity 53(2):231-244.
Lanata, José, Luis.
1993 Cambios Para Evolucionar: Las propiedades del registro arqueológico y la evolución de los grupos humanos en Patagonia y Tierra del Fuego, Trabajo presentado en las Segundas Jornadas de Arqueología de la Patagonia. Puerto Madryn.
1999 Notas Sobre Evolución y Arqueología , Hoy. ETNIA En prensa
Lanata, José, Luis., y Luis Alberto Borrero.
1994 Riesgo y Arqueología. En: Arqueología de Cazadores-Recolectores. Límites, Casos y Aperturas. J.L. Lanata & L.A Borrero comps, pp 129-143, Arqueología Contemporanea 5. Bs.As.
Lanata, José, Luis y Hector Neff
1998 Discutiendo Algunas Escalas de la Transmisión Cultural: Artefacto y Espacio. En 1ª Reunión de Teoría Arqueológica de Sudamérica. En Prensa
Lazzari, Marisa
1994 Materias Primas Líticas en la Falda Occidental del Aconquija . Hacia un Modelo Regional de Intercambio. Los Primeros Pasos V 1 AINA. BS. AS.
MacArthur, R., H, and E. Pianka R
1966 On Optimal Use of a Patchy Environment. American Naturalist 100:603-609.
Madsen, David, B, and Dave Schmitt N
1998 Mass Collecting and the Diet Breadth Model: A Great Basin Example. Journal of Archaeological Science 25:445-455.
Mandryk, Carole, A. S.
1993 Hunter-Gatherer Social Costs and the Nonviability of Submarginal Environments. Journal of Anthropological Research 49:39-71.
Markgraf, V.
1980 Paleoclimatic changes during the last 15,000 years in Subantartic an arid environments in Argentina (South America). Paper presented at The 5th. international Conference of Palinology, England.
1985 Paleoenvironmental History of the last 10,000 years in Northwestern Argentina. In Zbl. Geol. Palaont.
1987 Paleoclimates of the Southern Argentine Andes. Current Research in the Pleistocene
1989 Paleoclimates in Central and South America since 18,000 BP based on pollen and lake-level records. Quaternary Science Reviews, .
Margalef, Ramon
1980 Ecología. 2nd ed. Ediciones Omega, Barcelona.
Markgraf, V., and J. Bradbury P
1982 Holocene Climatic Stratigraphy of South America. Chronoestratigraphic Subdivision of the Holocene. Striae.
McGlone, M., S, A. Kershaw P, and Vera Markgraf
1993 El Niño/Southern Oscillation Climatic Variability in Australasian and South American Paleoenvironmental Records. In El Niño, Historical and Paleoclimatic Aspects of the Southern Oscillation, edited by H. Diaz F, and V. Markgraf, pp. 435-463. Cambridge University Press.
McNaughton, S., J, M. Oesterheld, and D. Frank A
1989 Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats. Nature 341:142-144.
McNaughton, S., J, O. Sala E, and M. Oesterheld
1993 Comparative Ecology of African and South American Arid to Subhumid Ecosystems. En Biological Relationships Between Africa and South America, edited by Goldblatt Peter, pp. 548-567. Yale University Press, New Haven.
Minnegal, Mónica
1997 A Necessary Unity: The Articulation of Ecological and Social Explanations of Behaviour. Journal of the Royal Anthropological Institute 2:141-158.
Muscio, Hernán, Juan.
1996 Poblamiento Humano y Evolución en la Puna Argentina - Desarrollo Teórico Para la Arqueología del Valle de San Antonio de Los Cobres, Salta. Tesis de Licenciatura en Cs. Antropológicas. UBA.
1998(a) Variabilidad de los Recursos Potenciales en la Puna: Desarrollo Teórico para su Aplicación en Arqueología Evolutiva. En Notas Gauterianas Nº 1, Publicación del Museo de Historia Natural Dr. Ricardo S Vadell.
1998(b) Patrones Espacio - Temporales de la Variabilidad Ambiental en la Puna Argentina: Algunas Implicancias Para la Ecología Humana Prehistórica del N.O.A y Para la Estructura Arqueológica Regional. Cuadernos del INA - En Prensa.
1998(c) Arqueología de San Antonio de Los Cobres (SAC), Poblamiento Prehistórico y Evolución en el N.O.A.: Primeras interpretaciones del registro arqueológico de superficie. MS
Nelson, Margaret, C
1996 Technological Strategies Responsive to Subsistence Stress, In: Evolving Complexity and Environmental Risk in the Prehistoric Southwest. Eds J. Tainter & B.B. Tainter, Addison-Wesley.
Nelson, Margaret, C, and Heidi Lippmeier
1993 Grindging-Tool Design as Conditioned by Land-Use Pattern. American Antiquity 58(2):286-305.
Nielsen, Axel
1995 El Pensamiento Tipológico Como Obstáculo Para la Arqueología de los Procesos de Evolución en Sociedades Sin Estado. comechingonia 8:21-46
Nuñez, Lautaro y Calogero M. Santoro
1988 Cazadores de la Puna Seca y Salada del Area Centro - Sur Andina (Norte de Chile). Estudios Atacameños 9 pp 11-60
Nuñez Lautaro, Grosjean Martin, Messerli Bruno; y Hans Scherelier 1996 Cambios Ambientales Holocénicos en la Puna de Atacama y sus Implicancias Paleoclimáticas. Estudios Atacameños Nº 12, 31-40
O'Connell, James, F
1994 Etnoarchaeology and Behavioral Ecology
Olivera, Daniel, y Dolores Elkin
1994 De Cazadores y Pastores: El Proceso de Domesticación de Camélidos en la Puna Meridional Argentina. Zooarqueología de Camélidos 1:95-124.
Pianka, E., R
1982 Ecología Evolutiva. Omega, Barcelona.
Pintar, Elizabeth
1992 Tool-Kits and Raw Material Use: Archaic Hunters of The Puna, Paper Presented at the 57th Anual Meeting of the Society for American Archaeology. Pittsburgh, April 1992.
1994 Movilidad, Artefactos y Materias Primas: La Organización Tecnológica en la Puna Desértica, Actas y Memorias del XI Congreso Nacional de Arqueología Argentina -San Rafael- Mendoza.
Rapoport, Eduardo, H, y Barbara Drausal
1975 Tácticos y Estrategas r, K y S O S. En Aerografía, Estrategias Geográficas De Las Especies, edited by E. Rapoport, pp. 111-128. FCE, México.
Ratto, Norma
1980 Elección de rocas y diseño de artefactos :propiedades físico - químicas de las materias primas líticas de ICc. 4. Presentado al XI Congreso de Arqueología de Chile, Santiago.
Sanguinetti de Bórmida, A.C., D.E. Curzio, E.A. Crivelli Montero y A. Chauvin,
1998. Asentamientos y tecnología de los cazadores-recolectores prehistóricos de El Manantial, Patagonia Septentrional Argentina. II Congreso Argentino de Americanistas, Tomo 2, pp. 437-459, Sociedad Argentina de Americanistas, Bs. As.
Schobinger, J.
1988 Prehistoria de Suramérica. 1988 ed. Alianza, Madrid.
Sherratt, Andrew
1997 Climatic Cicles and Behavioural Revolutions: The Emergence of Modern Humana and the Beginning of Farming. Antiquity 71:271-287.
Smith, Eric, Alden.
1983 Antropological Applications of Optimal Foraging Theory: A Critical Review. Current Anthropology 24(5).
1988 Risk and uncertainty in the original affluent society: evolutionary ecology of resource-sharing and land tenure. In Hunters and Gatherers 1. History, evolution, and social change, vol. 1, edited by Ingold T, Riches D, and Woodburn J, p. 222/251. Berg, Oxford.
Soriano, Alberto
1987 El Pastoreo Como Disturbio: Consecuencias Estructurales y Funcionales, Ecología 9:132-139.
Spielmann, K, A y Angstadt-Leto, Eric, A
1996 Hunting, Gathering and Helth in the Prehistoric Southwest. Evolving Complexity and Environmental Risk in the Prehistoric Southwest. 1st ed., edited by Santa Fe Institute, pp. 169-196. Addison-Wesley.
Steele James, Adams Jonathan and Sluckin Tim
1998 Modelling Paleoindian Dispersals World Archaeology Vol 30 (2):286-305
Stephens, D., W, y J. Krebs R
1986 Foraging Theory. Princeton: Univ.Press 1 Ed.
Tattersall Ian
1997 De Africa ¿una... y otra vez? Investigación y Ciencia Nº 249, Junio: 20-28
Tankersley, Kenneth, B
1994 The Effects of Stone and Technology on Fluted-Point Morphometry. American Antiquity 59(3):498-510.
1998 Variation in The Early Paleoindian Economies of Late Pleistocene Eastern North America. American Antiquity 63(1):7-20.
Torrence, Robin
1989 Tools as Optimal Solutions. In Time, Energy, and Stone Tools, edited by Robin Torrence, pp. 1-6. Cambridge University Press, Cambridge.
Vrba, Elizabeth, S
1996 Climate, Heterocrony, and Human Evolution. Journal of Anthropological Research 52:1-28.
Winterhalder, Bruce
1993 Work, Resources and Population in Foraging Societies. Man 28:321-340.
Wobst, Martin, H
1993 Mobility And Gene Flow: Some Biological and Social Thoughts on the Paleolithic.Essays in Honor of R..B. Woodbury, University of Massachusetts -Amherst.
Yacobaccio, Hugo, D, y Celina Madero M
1992 Zooarqueología de Huachichocana III (Jujuy, Argentina). Arqueología FFyL UBA 2:149-189.
Yacobaccio, Hugo, Daniel.
1983 Una Adaptación Regional en los Andes Centro-Sur. Relaciones de la Sociedad Argentina de Antropología (N.S.) XVI.
1985 Explotación Complementaria de Recursos en Sociedades Cazadoras-Recolectoras Surandinas, Cuadernos del INA V10.
1991 Sistemas de Asentamiento de los Cazadores-recolectores Tempranos de los Andes Centro-Sur. Tesis para optar al grado de Doctor en Filosofía, FFyL (UBA).
1994 Biomasa Animal y Consumo en el Pleistoceno-Holoceno Surandino. Arqueología 4 FFyL UBA
1998. The Evolution of South Andean Hunter-Gatherers. Proceedings of the XIII Congress of the International Union of Prehistoric and Protohistoric Sciences, Volume 5, pp. 389-394, A.B.A.C.O. Edizioni, Forli
Young, David, A, and Robert Bettinger L
1995 Simulating the Global Expansion in the Late Pleistocene. Journal of Archaeological Science 22. pp 89-92
NOTAS
- La estasis propuesta es un fenómeno resuelto en una escala temporal del orden de los 10 Ka, representado como la parte plana de una curva de evolución demográfica que comienza a crecer exponencialmente en torno a los 5 Ka BP. Su escala espacial mínimamente abarca el NOA (Muscio 1996).
- El concepto de Bioma refiere a una comunidad de animales y plantas que ocupan un área climáticamente uniforme en una escala continental (Hugget 1991)
- El ecotono es la transición espacial entre dos o más comunidades distintas, y puede tener una extensión lineal considerable (Butzer 1985, Hugget 1991)
- En los modelos de selección de tipo K y selección de tipo r , los procesos denso-dependientes son los que ocurren en un contexto de alta densidad de organismos en su ambiente. En tales contextos las poblaciones tipo K son aquellas en las que la supervivencia depende en mayor grado de la competencia. Contrariamente, en las poblaciones de tipo r la densidad de individuos y la competencia no son factores importantes en la supervivencia, siendo los relevantes la inestabilidad y las fluctuaciones ambientales (Pianka 1982).
- Un deme es una Población Local con un hábitat particular en el área de distribución de una especie.
- La adaptividad potencial es una medida de la probabilidad de una entidad evolutiva de ser replicada, lógicamente independiente del proceso de selección (O’Brien y Holland 1992). En este marco una variante adaptativa es un estado del rasgo (estructuras biológicas, conductas, artefactos) que aumenta la probabilidad de supervivencia y reproducción al portador; y una variante No Adaptativa del mismo rasgo es aquella que le disminuye esta misma probabilidad.
- Los nunamiut explotan un área anual de 63.700 Km2 con una distancia promedio por movimiento residencial de 69,5 Km, en un bioma con PP de 73/g/m2/yr, y alta estacionalidad (Kelly 1983)
- Sin embargo puede argumentarse, como hace Madsen (1993), que las presas pequeñas mediante una táctica de caza con redes o trampas -u otras similares- pueden mejorar el rendimiento. No obstante, esto implica una estrategia de mucho menor beneficio frente a la que logra efectivamente consumir, con una mayor frecuencia, a los artiodáctilos. Un caso contrario podría ser la caza masiva.
- Esta línea de argumentación tiene sustento empírico y teórico cuando se coteja el comportamiento de la Biomasa de Herbívoros de los diversos biomas terrestres en relación a la Biomasa Primaria (McNaughton et al 1989, Pianka 1982), muy especialmente en los paisajes áridos como el de la Puna (Mc Naughton et al 1993, y particularmente para los camélidos (Rabinovich 1989).
- La etología de los camélidos es muy flexible en sus patrones de movilidad, por lo que debe ser muy sensitiva de la degradación de la cobertura vegetal. Frente a la disminución de las pasturas en el hábitat los camélidos aumentan la movilidad migratoria, para disminuirla en ambientes más ricos. En el caso de la vicuña esta flexibilidad le permite explotar una mayor rango ambientes áridos (ver Flannery et al 1989).
- El valor obtenido de esta manera corresponde a la Utilidad Energética por Tiempo de Procesamiento (Tpi) el cual es una fracción de la Tasa de Calorías por Tiempo de Manejo. Luego resulta de : Tpi = Ei / tpi .
- La recolección masiva de los huevos de suri puede llevar a que este recurso aumente levemente su posición en el ranking si se realiza el cálculo a partir de otras premisas u observaciones, respecto a los tiempos de procesamiento.
- El producto µ está compuesto por dos magnitudes de energía, por espacio: hectárea (Ht) y por tiempo: horas (Hrs) . Se unidad resulta : Kcal2 / Ht.Hrs. Es apropiado transformar este producto en un índice, normalizándolo al valor 106 , lo que permite trabajar sin unidades y con un número de dígitos manejables.
- El número de combinaciones posibles de series de recursos con sus tres estados de disponibilidad en el paisaje resulta de 39. Si bien esto está asumiendo un estatus equiprobable para cada variable cuando quizás muchas covarían, advierte la alta variabilidad esperada de ambientes de escala local.
- El gráfico no muestra esta situación pero puede demostrarse fácilmente con un cociente donde la media de µ sea el numerador y la superficie del rango el denominador, aumentando el número de ambientes de la serie. En teoría el máximo de ambientes incluidos se alcanza cuando los costos de la movilidad interceptan el rendimiento promedio de toda la serie.
Buscar en esta seccion :