 Especial NAyA Volumen 1
Especial NAyA Volumen 1
PASTOREO Y MANEJO DE CAMELIDOS EN LA EPOCA LAMBAYEQUE : DATOS ZOOARQUEOLOGICOS
POR:
Ph.D. JONATHAN D. KENT (1)
Biól. VICTOR F. VASQUEZ SANCHEZ (2)
Arql. TERESA E. ROSALES THAM (2)
(1) Department of Sociology, Anthropology and Behavioral Sciences, Metropolitan State College of Denver, Denver, Colorado, 80217-3362. EE.UU. (kentj@mscd.edu).
(2) Centro de Investigaciones Arqueobiológicas y Paleoecológicas Andinas-"ARQUEOBIOS", Facultad de Ciencias Sociales, Sección de Post-grado, Universidad Nacional de Trujillo, La Libertad, Perú. (trosal@unitru.edu.pe).
INTRODUCCION
Avances recientes en el estudio de osteometría, de la morfología dentaria y de las fibras han permitido a los zooarqueólogos distinguir entre las cuatro formas existentes de camélidos con mayor certeza. Desafortunadamente, dichos estudios están siendo utilizado independientemente por la mayoría de zooarqueólogos. Esta situación tiene por consecuencia la falta de un nivel muy alto de confiabilidad en cada método por la carencia de verificación de un método por otro.
En 1990-1996, excavaciones en el sitio arqueológico de Huaca Cao Viejo revelaron entierros humanos prehistóricos con ofrendas de camélidos. Por lo común, partes del cuerpo de un solo camélido fueron puestos en las tumbas humanas. Incluidos como parte de dichas ofrendas fueron cráneos, huesos post-craneales, la piel y fibra de ejemplares individuales de camélidos. Este descubrimiento nos proveyó la oportunidad de comprobar los tres métodos de diferenciar entre las formas de camélido, aplicándolos a la misma muestra.
El sitio es un montículo de adobes construido durante la época Moche, entre el siglo I y el VIII d.C. y se ubica en la margen derecha de la parte baja del valle de Chicama en la costa norte del Perú, y está dentro de la zona de vida Desierto Desecado Subtropical.
Después de su abandono por la gente mochica, "...ocurrió un desmoronamiento masivo de las estructuras y la posterior utilización del sitio como cementerio durante la época Lambayeque." (Franco et al. 1994: 147).
De acuerdo a W. Alva (1985), la Cultura Lambayeque experimentó su época "clásica" durante los siglos VIII al XIII, pero tuvo su inicio desde los siglos VI y VII. Shimada (1985), denomina a esta manifestación arqueológica lambayecana como Sicán. Teniendo en cuenta que en la literatura del Valle de Chicama se usa más el término Lambayeque al referirse a esta cultura arqueológica, utilizaremos esta palabra en vez de Sicán.
Aunque en otros estudios de entierros lambayecanos se ha puesto mucha atención a rasgos como la posición del muerto, la presencia de máscaras y placas de cobre, estilos de cerámica, etc., pocas veces se ha llevado a cabo un estudio detallado de los restos orgánicos encontrados como ofrendas dentro de tumbas lambayecanas.
Un énfasis de nuestro análisis es en los restos de camélidos incluidos en las tumbas lambayecanas como ofrendas.
En algunas tumbas se han puesto partes del cuerpo de uno o más camélidos como ofrendas. Las partes constan comúnmente de las patas y del cráneo de un camélido. Esta práctica funeraria nos proveyó la oportunidad de comparar los métodos de identificación de restos de camélido al
nivel específico derivados de estudios del cráneo con métodos de identificación derivados de la osteometría de huesos post-craneales y ellos basados en el estudio de fibras.
METODOS
Para el desarrollo de la presente investigación, se procedió de la siguiente manera:
En cuanto a la selección de la muestra a estudiarse, se obtuvo una relación de la cantidad de entierros que pertenecen a la fase Lambayeque dentro de la parte del sitio excavado, lo cual contabiliza 259 entierros y a cada uno de ellos se les asignó un número de muestra.
Con la relación de entierros registrados, se realizó un muestreo aleatorio para seleccionar 50 de los entierros lambayecanos, según los procedimientos estadísticos no-paramétricos. Esta cantidad fue escogida para darnos una muestra representativa, confiable y sin sesgos. Decidimos optar para un error de menos de un sigma, con una confiabilidad de 95% .
En cuanto a la identificación de los restos de camélidos hasta el nivel de especie, se aprovecharon tanto métodos cualitativos como los cuantitativos. Con respeto a métodos cualitativos, utilizamos características tales como la forma del diente y la presencia de esmalte en la cara labial y lingual de los incisivos basándonos en los criterios de Wheeler (1982). Además, utilizamos una comparación visual microscópica de fibras con muestras de fibra controladas.
De acuerdo al estudio de Wheeler, incisivos de forma espatulada con esmalte en ambos lados pertenecen o a la llama o al guanaco. Incisivos de forma espatulada con esmalte solamente en la cara labial pertenecen a la alpaca, mientras incisivos de forma rectangular con esmalte solamente en la cara labial indican vicuñas.
Como algunas muestras arqueológicas estuvieron en buen grado de conservación y aún conservaron partes con piel y fibra, se extrajo una muestra de fibra para su análisis microscópico lo cual se llevó a cabo en Lima en el Laboratorio del Programa de Ovinos y Camélidos Americanos de la Universidad Nacional Agraria, La Molina. Vimos este tipo de análisis como muy importante para añadir otra variable útil en la identificación taxonómica, dado que las fibras pueden mostrar características diagnósticas que permiten diferenciar las cuatro especies de camélidos, como señala Reigadas (1994).
En la temporada de campo de 1990, una muestra de fibra de camélidos recuperada de entierros lambayecanos fue mandada al mismo Laboratorio para identificarse. Los resultados indicaron la presencia tanto de fibras de "llama" como fibras de "guanaco", por lo tanto decidimos mandar 12 muestras de fibra perteneciendo a nuestra muestra aleatoria, unas de patas y otras de cráneos. El protocolo utilizado, consiste en una comparación microscópica con una serie de muestras comparativas del laboratorio. Los diámetros de las fibras en la muestra fueron comparados con los diámetros de las fibras de la serie comparativa. Es de notar que la mayoría de las muestras comparativas se obtuvieron del pecho del animal, siendo ausentes muestras comparativas del cráneo y patas.
En cuanto a métodos cuantitativos, decidimos usar la osteometría según las variables identificadas por Kent (1982) como las más útiles para efectuar dicha identificación basándose en un análisis discriminante. También tomamos unas medidas utilizadas por Miller y Gill (1990). En total, se trata de un juego de 88 medidas del esqueleto postcraneal sugeridas por Kent, más 9 medidas del astrágalo y calcáneo utilizadas por Miller y sus colegas. Además decidimos tomar una serie de medidas del cráneo y la mandíbula para que posteriormente podamos tener datos comparativos de la población de cada especie identificada y para comparar con las tomadas por otros investigadores como Herre (1952). Las medidas craneales no fueron utilizadas en la identificación de camélidos al nivel específico por la falta de pautas estadísticas para efectuar tal identificación. Las medidas se efectuaron con un vernier de ±0.05 mm. de precisión, realizando tres veces las medidas y sacando un promedio de ellas para evitar sesgos. Las fórmulas del análisis discriminante fueron puestas en una hoja de cálculo Lotus y los datos osteométricos fueron ingresados a ella.
Comparación de métodos
La meta de este estudio es comparar los datos de identificación específica obtenidos por los tres métodos y era menester tener en cada tumba partes del cuerpo y del cráneo del mismo animal. Tenemos tres razones para concluir que muy probablemente los cráneos y las patas incluidos como ofrendas en cada tumba puedan aparearse: 1) se encuentran dentro del mismo contexto y por consecuencia representan un solo momento de deposición; 2) en cuanto a la fibra que cubre las patas, se nota que los colores y los patrones de colores que ellos forman son los mismos que los que se encuentran cubriendo el cráneo y la mandíbula; 3) los datos osteométricos indican que en la mayoría de los casos, las patas pertenecen a la misma especie que nos hemos desprendido para la mandíbula en base a la morfología dentaria.
Aunque no es el énfasis de esta ponencia, tomamos en cuenta datos sobre la edad y el sexo del animal utilizando características tales como el desarrollo, erupción y desgaste de los dientes, fusión de las epífisis de los huesos largos, clausura de súturas craneales y el tamaño de los caninos.
Es de notar que hay una falta de acuerdo en los estudios publicados entre las estimaciones de edad basadas en desgaste dentario. Esta puede deberse a una diferencia entre los pastos disponibles y consumidos por los camélidos estudiados en Chile por Puig y Monge (1983) y en la Sierra Sur del Perú por Kent y Wheeler. Otra alternativa es un manejo diferente por el ser humano en cuanto a pastoreo de estos camélidos.
Otra indicación del manejo son las anomalías dentales y/o óseas, que podían producirse por una crianza cuyo resultado sería la alteración del genoma del animal. Si este fuera el caso, probablemente se trata de animales domesticados en vez de un animales silvestres.
RESULTADOS
Hay 15 entierros de los 50 estudiados que si tienen camélidos como ofrendas (30%). Con dos excepciones, los entierros que tienen camélidos se han cavado dentro del relleno y adobes cubriendo el montículo y se depositó el difunto.
De los 21 ejemplares de camélido en estos 15 entierros, 13 de ellos (61.9%) fueron identificados al nivel específico utilizando la morfología de los incisivos u osteometría o una combinación de estos métodos.
De estos 13 ejemplares, la morfología dentaria fue el único dato para efectuar la identificación en 3 de los casos (el grupo "A" del cuadro Nº 1), siendo identificados "llama" o "guanaco" para 2 (Muestra 17, Individuo I y Muestra 129, Individuo I) y "alpaca" para uno (Muestra 104, Individuo I).
La osteometría fue el único criterio para la identificación específica de un ejemplar (grupo "B" del cuadro Nº 1; Muestra 200, Individuo II). En este caso, las medidas resultan en la identificación de "llama" por algunos huesos y la identificación de "guanaco" por otros. Nuestra interpretación de esta mezcla es que se trata de un posible híbrido.
Cuadro Nº 1.- Identificaciones de las especies de Camelidae en Entierros Lambayecanos de la Huaca Cao Viejo-Costa Norte del Perú.
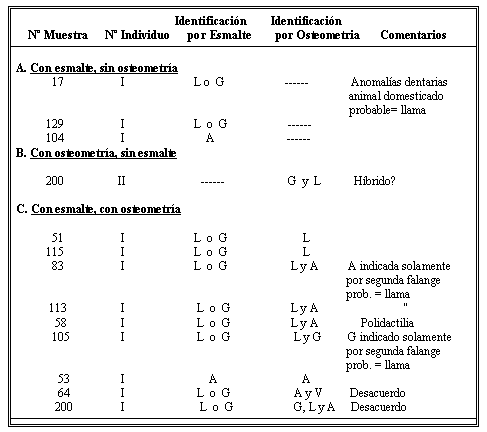
Para los 9 ejemplares restantes (grupo "C" del cuadro Nº 1), podemos contar con ambos métodos para efectuar la identificación específica. En dos de ellos (Muestra 51, Individuo I y Muestra 115, Individuo I), la morfología dentaria indica "llama" o "guanaco" mientras la osteometría indica "llama" para todos los huesos medidos. Nuestra interpretación es que se trata de una "llama".
En tres casos, la dentición indica "llama" o "guanaco", mientras las medidas indican una "llama" para unos huesos y una "alpaca" para otros. En esta categoría se nota que en dos de los tres (Muestra 83, Individuo I y Muestra 113, Individuo I), el único hueso cuyas medidas indican "alpaca" es la segunda falange, mientras las otras medidas indican "llama". Por eso, concluimos que hay problemas con la formula discriminante para este hueso y que la identificación correcta sería "llama". En el tercer caso (Muestra 58, Individuo I), se trata de un animal con una anomalía genética, polidactilia. Es de presumir que medidas de un animal afectado de tal manera no son confiables. Al mismo tiempo, dicha anomalía es más frecuente en animales domesticados que en animales silvestres. Por eso, nos desprendemos que se trata de una "llama".
Hay un ejemplar (Muestra 105, Individuo I) caracterizado por morfología dentaria de una "llama" o de un "guanaco" y cuyas medidas sugieren una "llama" por algunos huesos y un "guanaco" por otros. En este caso es una vez más la segunda falange que nos da la clasificación de "guanaco", subrayando por los tanto el problema con el uso de este hueso para efectuar la identificación correcta.
Se encuentran solamente un ejemplar cuya identificación por ambos métodos correspondía a "alpaca" (Muestra 53, Individuo I). Otra vez más tenemos un acuerdo entre los dos métodos.
Al fin, hay dos ejemplares mostrando desacuerdos serios entre la identificación por morfología dentaria y osteometría. En el primer ejemplar (Muestra 64, Individuo I), hay esmalte en ambos lados de los incisivos espatulados, indicando una "llama" o un "guanaco" de acuerdo a los criterios de Wheeler. Hemos podido tomar medidas de varios huesos postcraneales, entonces para las primeras falanges, la clasificación más probable indica una "alpaca", igual para otras medidas. En cuanto a las segundas falanges, la clasificación más probable cae entre una "alpaca" y una "llama", pero como mencionamos anteriormente, no tenemos mucha confianza en las clasificaciones por este hueso.
En el segundo caso (Muestra 200, Individuo I), la morfología dentaria indica una "llama" o un "guanaco". La osteometría indica un "guanaco" para algunos huesos de este individuo. Estos resultados encajan bien con la asignación derivada en base a los dientes. Sin embargo, para otros huesos, incluyendo algunas de las segundas y terceras falanges, tenemos como resultado la asignación a "llama". Finalmente, para un hueso, tenemos como resultado la asignación a "alpaca".
SUMARIO Y COMENTARIOS ADICIONALES
En términos generales, vemos un acuerdo bueno entre los datos para una identificación por la osteometría y por la morfología de los incisivos. En varios casos, el desacuerdo existente se deriva de las medidas de la segunda falange y no aconsejamos su uso futuro para este motivo.
Pero, tenemos que buscar un porque para los dos casos de desacuerdos.
Existen unas regularidades en estos dos casos:
1. Ambos casos de desacuerdo ocurren cuando hay esmalte en ambos lados de los incisivos espatulados, lo cual de acuerdo a Wheeler indicaría o una "llama" o un "guanaco"; y
2. La osteometría respectiva indica que algunos huesos son de una "alpaca".
Hay varias explicaciones posibles:
a. Es posible que el criterio de morfología dentaria siempre nos lleva a una identificación correcta pero la osteometría no siempre nos lleva a una identificación correcta por tener un margen de error;
b. También es posible que el criterio de morfología dentaria siempre nos lleva a una identificación correcta y que la osteometría no siempre nos lleva a una identificación correcta por haber sido derivado de animales de otras regiones, y que los casos de desacuerdos son animales con proporciones o tamaños diferentes de los que se utilizaron para derivar las formulas de clasificación;
c. Una tercera alternativa es que la osteometría nos lleva a una identificación correcta pero en algunos casos, el criterio de morfología dentaria tiene excepciones.
En cuanto a la primera alternativa, es cierto que para algunos huesos, la aplicación de las fórmulas de clasificación a la muestra original conllevó a una mala clasificación de un porcentaje de los animales medidos. Nuestros porcentajes de clasificación en el peor de los casos va de 75%(para la tercera falange) y en el mejor de los casos hasta un 100% para las primeras falanges.
Si asumimos para esta evaluación que podemos confiar mucho en los coeficientes de clasificación de las primeras falanges delanteras, vemos que existe en la muestra de la Huaca Cao Viejo un desacuerdo solamente en el caso de Muestra Nº 64, Individuo I. Vemos que este animal está clasificado osteometricamente como una "alpaca" para casi todos los huesos postcraneales, mientras la morfología dentaria nos dice que pertenece al grupo grande de camélidos (guanaco o llama). Si tenemos en cuenta los márgenes de error bajos para cada hueso, la probabilidad que
exista un error simultáneo para todos los coeficientes de clasificación de todos los huesos sería el producto de la probabilidad de una mala clasificación para cada hueso. En este caso, tendríamos tal error por casualidad una vez en 40'000,000. Por lo tanto, lo vemos difícil creer que todas los coeficientes de clasificación están erróneos para el mismo animal y de la misma manera por casualidad. En conclusión, rechazamos la primera alternativa.
En cuanto a la segunda alternativa, sería muy difícil decidir si los rasgos de camélidos de la zona de la costa norte del Perú difieren mucho o poco de los camélidos de otros lugares. En primer término, aunque es probable que los camélidos de la época Lambayeque pastaron con plantas disponibles localmente, no sabemos el efecto de ello sobre su biología. En segundo lugar, no hay camélidos en la costa norte actualmente que nos permitan obtener los datos comparativos. Por lo tanto, tenemos que trabajar con esta alternativa como una no rechazable.
La tercera alternativa está más factible de tratar. Sabemos que hay márgenes de error en los coeficientes de clasificación para muchas partes anatómicas, pero están conocidas y se pueden precisarlas. Además, Kent menciona que en algunos casos en la muestra de La Raya estudiada tanto por él como por Wheeler y por Miller, hay "alpacas" con esmalte en ambos lados de los incisivos, una observación que no había sido tomado en cuenta por Wheeler. Por lo tanto el criterio de clasificación taxonómica por la dentición no sería un criterio infalible para todas las clasificaciones.
Datos de análisis de fibras y su comparación con otros datos.
Como mencionamos anteriormente, un análisis de fibras procedentes de entierros lambayecanos excavados durante la temporada de 1990 proveyó la identificación de fibras tanto de "llama" como de "guanaco". Al recibir de las identificaciones de fibras que mandamos al mismo laboratorio en 1996, seleccionadas de nuestra muestra aleatoria, nos quedamos sorprendidos por los resultados. Todas las muestras se identificaron como fibras de "alpacas". Este resultado representa un desacuerdo serio con respeto a las identificaciones obtenidas por la dentición y la osteometría.
Dado el acuerdo generalmente bueno entre las identificaciones por la dentición y ellas obtenidas por osteometría, nos desprendemos que el desacuerdo radica principalmente en las identificaciones de las muestras de fibra. Las muestras que mandamos procedieron de las orejas, patas y cráneos de animales en las tumbas, mientras que los especímenes disponibles para comparación en el Laboratorio de Fibras Textiles procedieron principalmente del pecho y lomo de animales. Siendo que nuestra muestra fue seleccionada de partes del cuerpo del animal que no estuvieron en la muestra comparativa en el Laboratorio, es posible que su identificación fuese errónea debida a variaciones de las características de las fibras a través del cuerpo del animal,
aunque más adelante hacemos una evaluación de estos resultados en base a datos comparativos modernos de fibras de híbridos. Es de notar que en su estudio sobre las características de fibras de
camélidos, Reigadas(1994) subrayó la mayor variación en las fibras de las patas que en el lomo o pecho. Datos sobre las fibras de las orejas y los cráneos no fueron ubicados.
Para resumir, de los 15 entierros con camélidos, hemos identificado los taxones siguientes:
1. Camelidae
2. Lama sp. (llama, alpaca o guanaco)
3. Lama cf. glama, cf. guanicöe (o llama o guanaco [=camélidos grandes])
4. Lama glama (llama)
5. Lama pacos (alpaca).
En cuanto al número 3, los camélidos grandes, no podemos rechazar la posibilidad que sí habían "guanacos" como ofrendas en las tumbas. En primer lugar, unos de los coeficientes de clasificación de los huesos postcraneales dieron como resultado la asignación del hueso a esta especie. En segundo lugar, datos procedentes de unas muestras de fibra analizadas en 1990 por el personal del Laboratorio, señalan que unas de las muestras pertenecen a "guanacos". Por lo menos, tenemos que acercarnos a las identificaciones tomando en cuenta esta posibilidad.
La presencia de "llamas", "alpacas" y posibles híbridos dentro de las tumbas lambayecanas indica una manera de pastoreo enfocado en un rebaño mixto. Además, la presencia de un animal cuya edad fue alrededor de 3 meses (Muestra 104, Individuo I) sugiere que al menos unos entierros tuvieron lugar durante los meses de marzo a mayo, o sea al fin de la estación de lluvias en la sierra en los meses iniciales del invierno.
Crianza y Pastoreo en la costa Nor-Peruana
La pregunta que por mucho tiempo se han hecho los investigadores es que si hubo una crianza en ecosistemas costeros prehispanicos, o si los restos de camelidos encontrados en los depositos de la costa son o resultado de una crianza local o producto de un intercambio con pastores de zonas altoandina( ver por ejemplo, Shimada y Shimada 1985). Las anomalías genéticas como Polidactilia (Muestra 58), lesiones poróticas en las ampollas auditivas(Muestra 51) y mucha variación en la secuencia de erupción de los dientes y patrones de desgaste en los dientes, indicaría la posibilidad mas fuerte de un manejo local de rebaños por pastores lambayecanos. Aquí hay que enfatizar los datos de crianza y manejo de rebaños que realizan en la puna de Jujuy (Argentina) donde el cruce entre una llama por alpaca, da una descendencia F1 con características fenotipicas muy cercanas a la "llama", y características alpacunas como un tipo de fibra superior en finura incluso a la misma "alpaca"(Dr. Daniel Olivera, Com. personal, Julio 1997). Esto puede explicar el porque los análisis de fibras arqueológicas dieron como resultado a todas las muestras que proceden de "alpacas". Si analizamos detenidamente el dato moderno de los rebaños mixtos en la puna de Argentina, vemos que este modelo estaría explicando perfectamente la presencia de
rebaños mixtos y de los híbridos para la época lambayeque en un ecosistema costero prehispánico, la explicación del porque es sencilla: un rebaño mixto permite mantener la variabilidad del pool de genes, el vigor híbrido permite una mejor adaptación a otros ecosistemas, y finalmente se tiene a la mano carne, fibra de buena calidad y transporte.
REFERENCIAS
ALVA A., Walter
1985 Tempranas manifestaciones culturales en la región de Lambayeque. En Presencia Histórica de Lambayeque, editado por Eric Mendoza Samillán y otros, pp. 53-75. Ediciones y Representaciones H. Falconi, E.I.R.L., Lambayeque, Chiclayo.
FRANCO J., Régulo, César Gálvez M. y Segundo Vásquez S.
1993 Proyecto Arqueológico "El Brujo": Programa 1993 (Informe Final). Convenio: U.N.T-Fundación A.N.Wiese-IRC/LL. Informe archivado en la Fac. de Ciencias Sociales, Universidad Nacional de Trujillo y el Instituto Regional de Cultura, La Libertad, Trujillo.
HERRE, Wolf
1952 Studien über die wilden und domestizierten Tylopoden sudamerikas. Zoologische Garten, Leipzig, NF (19), Heft 2/4: 70-98.
KENT, Jonathan D.
1982 The Domestication and Exploitation of the South American Camelids:Methods of Analysis and Their Application to Circum-Lacustrine Archaeological Sites in Bolivia and Perú. PhD. Dissertation, Washington University-St. Louis. University Microfilms, Ann Arbor.
MILLER, George R. y Anne L. GILL
1990 Zooarchaeology at Pirincay, a Formative Period Site in Highland Ecuador. Journal of Field Archaeology, 17:49-68.
PUIG, Silvia y Susana MONGE
1983 Determinación de la edad en Lama guanicöe (Müller). DESERTA 7: 246-270. Mendoza, Argentina.
REIGADAS, María del Carmen
1994 Caracterizacíon de tipos de camélidos domésticos actuales para el
estudio de fibras arqueológicas en tiempos de transición y consolidación de
la domesticación animal. En: Zooarqueología de Camelidos: Perspectivas
Teóricas y Metodológicas (1º Parte) Año 1 Volumen 1, Grupo de Zooarqueología
de Camelidos, 1era. Edición, Buenos Aires-Argentina
SHIMADA, Izumi
1985 La cultura Sicán, caracterización arqueológica. En Presencia Histórica de Lambayeque, editado por Eric Mendoza Samillán y otros, pp. 76-183. Ediciones y Representaciones H. Falconi, E.I.R.L. Lambayeque, Chiclayo.
SHIMADA, Melody and Izumi Shimada
1985 "Prehistoric Llama breeding and herding on the North Coast of Perú". American Antiquity 50(1) 3-26.
WHEELER, Jane C.
1982 Aging Llamas and Alpacas by their Teeth. Llama World 1:12-17. Denver, Colorado.
Buscar en esta seccion :